It would thus seem that every phylogenetic analysis, in order to be valid, must include every species of bird [...], which is clearly impossible.
Harshman 2007:2
1. Celoptačí fylogeneze: sen, který došel naplnění
Fylogeneze ptáků je stále kontroverzním tématem, kde nový výzkum může mnoho změnit a přinést spoustu překvapujících závěrů – viz i studie, které o tomto tématu pravidelně zpracovávám pro blog. Až dosud jsme ale museli volit mezi studiemi, které téměř kompletně samplovaly malé skupiny (např. Bertelli & Porzecanski 2004, kteří analyzovali všechny popsané tinamy), nebo naopak studiemi s taxonomickým vzorkem obsahujících jen pár desítek "druhů" roztroušených napříč celým korunním kladem ptáků (Hackett et al. 2008; McCormack et al. 2012). Pokud jsme chtěli oboje najednou, museli jsme spoléhat na projekty entuziastů mimo primární literaturu – třeba výborný web Taxonomy in Flux Johna Boyda. Už několik let zpátky přitom Goloboff et al. (2009) úspěšně provedli analýzu čitající stěží uvěřitelných 73 060 taxonů: do takového množství by se pohodlně vešly všechny známé "druhy" žijících ptáků, ať už pod biologickým konceptem (pak je jich asi 10 000), nebo fylogenetickým konceptem (25 000 až 30 000). Otázkou času tedy bylo, než se o něco takového někdo skutečně pokusí, a asi měsíc zpátky se začaly po webu šířit informace, že takový projekt se už brzo dočká publikace od skupiny autorů pod vedením Waltera Jetze (viz i zmínku v nedávném článku).
Fylogeneze ptáků je stále kontroverzním tématem, kde nový výzkum může mnoho změnit a přinést spoustu překvapujících závěrů – viz i studie, které o tomto tématu pravidelně zpracovávám pro blog. Až dosud jsme ale museli volit mezi studiemi, které téměř kompletně samplovaly malé skupiny (např. Bertelli & Porzecanski 2004, kteří analyzovali všechny popsané tinamy), nebo naopak studiemi s taxonomickým vzorkem obsahujících jen pár desítek "druhů" roztroušených napříč celým korunním kladem ptáků (Hackett et al. 2008; McCormack et al. 2012). Pokud jsme chtěli oboje najednou, museli jsme spoléhat na projekty entuziastů mimo primární literaturu – třeba výborný web Taxonomy in Flux Johna Boyda. Už několik let zpátky přitom Goloboff et al. (2009) úspěšně provedli analýzu čitající stěží uvěřitelných 73 060 taxonů: do takového množství by se pohodlně vešly všechny známé "druhy" žijících ptáků, ať už pod biologickým konceptem (pak je jich asi 10 000), nebo fylogenetickým konceptem (25 000 až 30 000). Otázkou času tedy bylo, než se o něco takového někdo skutečně pokusí, a asi měsíc zpátky se začaly po webu šířit informace, že takový projekt se už brzo dočká publikace od skupiny autorů pod vedením Waltera Jetze (viz i zmínku v nedávném článku).
Nyní Jetz et al. (2012) svou práci konečně publikovali.
2. Co nám tempa diverzifikace a rapidní radiace říkají o vzorcích v ptačí diverzitě
2. Co nám tempa diverzifikace a rapidní radiace říkají o vzorcích v ptačí diverzitě
V první řadě je nutné zdůraznit, že kompletní ptačí fylogeneze je spíš třešničkou na dortu nové práce – lépe řečeno, je v ní obsažená proto, aby pomohla zodpovědět jiné, dost možná zajímavější otázky. Jetzovi a spol. jde hlavně o vzorce rozpoznatelné v ptačí diverzitě, a to jak v čase, tak i v prostoru. To vyžaduje syntézu makroekologických a fylogenetických přístupů. Dnešní pokročilé statistické modelování umožňuje vytřískat z molekulárních dat neuvěřitelné množství informací, od přiřazování absolutních časů k jednotlivým fylogenetickým uzlům (viz např. Brown & van Tuinen 2011) až po odhadování tempa diverzifikace a jeho změn (viz třeba můj článek o vangách, kde se povedlo odhalit období zvýšené speciace a najít pro ně věrohodné příčiny). Krásnou ukázkou toho jsou např. Alfaro et al. (2009), kteří ukázali, že ptačí klad Neoaves je produktem druhé nejmasivnější evoluční radiace v historii obratlovců hned po perkomorfních rybách, nebo Rabosky & Lovette (2008a), kterým se na severoamerických lesňáčkovitých (Parulidae) podařilo doložit hustotně-závislou diverzifikaci, v níž model tempo speciace závisí na počtu druhů, nikoli na čase (viz Stadler 2011).
Jsou to právě nekompletní vzorky (jak fylogenetické, tak i geografické), na nichž byl dosavadní výzkum u ptáků prováděn, který znemožňoval zhodnotit vliv rychlých diverzifikací a význam jejich míst původu na dnešní globální biodiverzitu ptáků. Nejobsáhlejší zatím dostupný dataset představili Sibley & Ahlquist (1990) se svou fylogenezí založenou na DNA-DNA hybridizaci, který k analýze ptačí rozmanitosti využili třeba Nee et al. (1992) – jeden z autorů této studie, Arne Mooers, se podílí i na Jetz et al. (2012). Sibleyho a Ahlquistova "tapisérie" však obsahovala pouhých 1700 druhů z celkových asi 10 000 a její topologie navíc ostře odporuje fylogenomickým studiím posledních let (Ericson et al. 2006; Hackett et al. 2008; Suh et al. 2011). Použitá distanční metoda nedokázala správně odkrýt dokonce ani základní dichotomii moderních ptáků na Palaeognathae a Neognathae, nemluvě už o bizarních pozicích (namátkou) perepelů, pěvců nebo kondorů. Studie založené na této tapisérii pak většinou naznačovaly, že ptáci prodělávali rychlejší diverzifikační události v tropech a v Jižní Americe ve srovnání se Severní Amerikou (viz např. Ricklefs [2006] u pěvců). Rozpoznání obecných vzorců také komplikují adaptivní radiace považované za mimořádné: typickými ukázkami jsou třeba galapážské pěnkavky (Grant & Grant 2008), madagaskarské vangy (Jønsson et al. 2012; Reddy et al. 2012 – viz příslušný článek), pěnicovití Starého světa aneb "Sylviidae" sensu lato (umělá skupina, jejíž zástupci jsou roztroušení napříč větší částí kladu Sylvioidea; viz Boyd 2012), výše zmínění lesňáčkovití, nebo kruhoočkovití (Zosteropidae), kteří odštěpují nové druhy šíleným tempem přesahujícím 2 druhy za milion let, jedním z nejvyšších mezi obratlovci, a to na celé ploše svého výskytu, zahrnující celé tropy Starého světa (Moyle et al. 2009a). Právě takovéto rapidně speciující klady Jetz et al. (2012) uvádějí coby příklad jevu, který nebyl plně vysvětlen a jeho implikace plně doceněny. Frekvence změn tempa diverzifikace a vliv jeho lokálních odchylek u malých kladů typu kruhooček ve srovnání s variacemi jdoucími napříč celým fylogenetickým stromem patří mezi nedostatečně prozkoumaná témata, a právě to se Jetz et al. (2012) snaží změnit.

3. Odhad fylogeneze: gory details
Zde právě přichází na řadu "kompletní" fylogeneze moderních ptáků. Autoři vytvořili fylogenetický strom čítající 9993 jednotlivých taxonů, a i když ne všechny zdroje by souhlasily, že je skutečně stoprocentně kompletní – Clements et al. [2012] např. odhadují počet ptačích druhů na 10 240 –, nemůže být sporu o tom, že rozsah díla je zcela bezprecedentní. Přirozeně se nabízí otázky, jak autoři dokázali "celoptačí" strom sestavit, když jsou genetická data k dispozici pouze od 6663 druhů (jak uvádějí sami autoři) – morfologie samozřejmě nic neřeší, protože nakódovat informativní znaky pro několik tisícovek taxonů by vyžadovalo strávit v muzejních depozitářích několik století – a když i daleko menší analýzy často staví současné počítače před velké problémy (Hackett et al. 2008). Sekce "Metody" v samotném textu studie postup objasňuje jen povrchně; to hlavní se nachází až v doplňkovém online materiálu. Tam autoři svou neradostnou situaci dále přibližují: software implementující sofistikované metody analýzy (např. BEAST, jednička v relaxovaných molekulárních hodinách, nebo MrBayes 3.2 a jeho působivé schopnosti v oblasti volby substitučních modelů) nezvládá odhadovat stromy s více než 1000 taxony, zatímco metody, které takto omezené nejsou (třeba PATHd8, vyhlazující tempo evoluce), trpí mnoha nedostatky: omezují tvar priorních rozdělení dob divergence okolo kalibračních fosilií a poskytují zavádějící odhady stáří uzlů. Autoři postupovali tak, že zkombinovali základní "kostru" (časově kalibrovaný strom velkoškálových příbuzenských vztahů v rámci ptáků) se samostatně odhadnutými fylogenezemi pro menší klady.
Topologie této kostry je založena na fylogenomických studiích ptáků jako celku (Ericson et al. 2006; Hackett et al. 2008) a pěvců (např. Barker et al. 2004; Jønsson et al. 2008). Zahrnuje celkem 158 koncových taxonů a 245 (u kostry odvozené z Hackett et al. 2008) nebo 249 (z Ericson et al. 2006) vnitřních uzlů, z nichž některé (ty s bootstrapem nad 70% nebo posteriorní pravděpodobnostní nad 0,95) si autoři vynutili po vlastní analýze (204 u Hackett et al., 212 u Ericson et al.). Autoři v tomto místě diskutují přetrvávající kontroverze ohledně dělení Neoaves na klady Metaves a Coronaves, které – jak je dobře známo – stojí a padá se zahrnutím 7. intronu β-fibrinogenu (Ericson et al. 2006; Hackett et al. 2008; Morgan-Richards et al. 2008). Signál podporující monofylii Metaves může být pouhým artefaktem špatně zarovnatelných dlouhých indelů, které jsou pro tento lokus typické (Morgan-Richards et al. 2008). Právě proto autoři důsledně používají dvě topologické kostry: ta, která je odvozená z Hackett et al. (2008), je založená na 19 lokusech a dichotomie Metaves/Coronaves na ní přítomná je; ta z Ericson et al. (2006: ESM-Figure 6) stojí na 4 genech (bez β-fibrinogenu) a Metaves ani Coronaves na ní nejsou. Topologie pěvců na obou podkladových stromech se řídí spíše jinými studiemi, hlavně pak Barker et al. (2004).
Ze 158 koncových taxonů 29 zahrnuje 4 a méně druhů, 129 naopak 5 a více (největší z nich dokonce plných 460). Každý z 9993 rozeznávaných druhů – autoři svůj výběr druhů odvozovali především ze světového seznamu Birdlife V3 z června 2010 – byl potom na základě předchozích fylogenetických studií zařazen do jednoho z těchto koncových taxonů. Následně Jetz et al. (2012) provedli fylogenetickou analýzu nad všemi druhy z jednoho koncového taxonu, pro které jsou dostupná genetická data, a celý postup opakovali, dokud nezískali fylogeneze pro všech 129 korunních kladů s více než 4 druhy. Při těchto analýzách si nevynucovali žádná omezení (constraints); data zde měla plnou svobodu diktovat výsledky. Každý z těchto 129 dílčích stromů byl také datován metodou relaxovaných molekulárních hodin.
Následně Jetz a spol. přistoupili k analýze 3323 taxonů, pro něž genetická data k dispozici nejsou. V tomto případě předpokládali, že taxonomie aproximuje fylogenezi: 2997 druhů bez dostupných DNA sekvencí je řazeno do "rodů", od nichž genetická data alespoň jeden druh máme. V takovém případě autoři prostě problematické taxony přilípli k jejich lépe prozkoumaným bratrancům. Tento postup má ale evidentní nevýhodu: předpokládá monofylii "rodů", přestože víme, že ptačí "rody" monofyletické velice často nejsou (Harshman 2007) a v případě pěvců mohou být dokonce druhy z jednoho "rodu" rozptýleny napříč několika "čeleděmi" (tamtéž). Jetz et al. (2012) proto sáhli po kompromisu: příslušenství k "rodu" ignorovali, pokud se ukázal být nonmonofyletickým na základě genetických dat, a dali druhu možnost zaujmout jakoukoli pozici uvnitř nejmenšího kladu obsahujícího všechny zástupce daného "rodu". Genetická data nejsou vůbec dostupná pro 214 "rodů" s celkem 326 druhy; tam se autoři spoléhali na zařazení do větších skupin. Logická otázka zní, jak vůbec do stromu zakomponovat taxony, pro něž nemáme žádná data (s výjimkou vágních priorů typu "X patří někam sem"). Zde se opět projevuje výhoda bayesovského přístupu: v případě absence sekvenčních dat Jetz et al. (2012) prostě nechali Markovovy řetězce Monte Carlo (MCMC) samplovat stromy z birth-death priorů (které jsou výhodné tím, že je lze kombinovat s "constraints" typu monofyletických "rodů"), přičemž tempo vymírání ("deaths" v birth-death modelu) bylo nastaveno na nulu. Není to proto, že by snad autoři věřili, že ptáci během své evoluci nevymírali, ale kvůli studiím ukazujícím, že homogenní tempo extinkce nezanechává žádný typický vzorec ve tvaru stromů.
Jsou to právě nekompletní vzorky (jak fylogenetické, tak i geografické), na nichž byl dosavadní výzkum u ptáků prováděn, který znemožňoval zhodnotit vliv rychlých diverzifikací a význam jejich míst původu na dnešní globální biodiverzitu ptáků. Nejobsáhlejší zatím dostupný dataset představili Sibley & Ahlquist (1990) se svou fylogenezí založenou na DNA-DNA hybridizaci, který k analýze ptačí rozmanitosti využili třeba Nee et al. (1992) – jeden z autorů této studie, Arne Mooers, se podílí i na Jetz et al. (2012). Sibleyho a Ahlquistova "tapisérie" však obsahovala pouhých 1700 druhů z celkových asi 10 000 a její topologie navíc ostře odporuje fylogenomickým studiím posledních let (Ericson et al. 2006; Hackett et al. 2008; Suh et al. 2011). Použitá distanční metoda nedokázala správně odkrýt dokonce ani základní dichotomii moderních ptáků na Palaeognathae a Neognathae, nemluvě už o bizarních pozicích (namátkou) perepelů, pěvců nebo kondorů. Studie založené na této tapisérii pak většinou naznačovaly, že ptáci prodělávali rychlejší diverzifikační události v tropech a v Jižní Americe ve srovnání se Severní Amerikou (viz např. Ricklefs [2006] u pěvců). Rozpoznání obecných vzorců také komplikují adaptivní radiace považované za mimořádné: typickými ukázkami jsou třeba galapážské pěnkavky (Grant & Grant 2008), madagaskarské vangy (Jønsson et al. 2012; Reddy et al. 2012 – viz příslušný článek), pěnicovití Starého světa aneb "Sylviidae" sensu lato (umělá skupina, jejíž zástupci jsou roztroušení napříč větší částí kladu Sylvioidea; viz Boyd 2012), výše zmínění lesňáčkovití, nebo kruhoočkovití (Zosteropidae), kteří odštěpují nové druhy šíleným tempem přesahujícím 2 druhy za milion let, jedním z nejvyšších mezi obratlovci, a to na celé ploše svého výskytu, zahrnující celé tropy Starého světa (Moyle et al. 2009a). Právě takovéto rapidně speciující klady Jetz et al. (2012) uvádějí coby příklad jevu, který nebyl plně vysvětlen a jeho implikace plně doceněny. Frekvence změn tempa diverzifikace a vliv jeho lokálních odchylek u malých kladů typu kruhooček ve srovnání s variacemi jdoucími napříč celým fylogenetickým stromem patří mezi nedostatečně prozkoumaná témata, a právě to se Jetz et al. (2012) snaží změnit.

Fylogeneze všech 9993 druhů ptáků. Uzly, na nichž byl aspoň pro 25% testovaných strom odhalen posun k vyšší rychlosti diverzifikace, jsou označeny koláčovými grafy, kde černá barva označuje podíl stromů, kde byl posun optimizován přímo na tento uzel, a šedá barva podíl stromů, kde k posunu došlo teprve uvnitř tohoto uzlu. Posuny jsou počítány jen jednou, takže např. posun u uzlu B se nezapočítává do šedé oblasti na uzlu A. Počet zaznamenaných posunů může být příliš nízký, protože kombinace vnořených posunů se často nepovede identifikovat na jednom stromě. Společné posuny A+B, H+I, J+L a N+O se např. vyskytují na méně než 5% stromů, ale jeden z dvojice už na 80%. S počtem vnořených posunů se efekt zhoršuje: J+L+Q se pohromadě nevyskytují ani na jednom stromě, ale aspoň jeden z nich už na víc než 96% stromů. Některé série vnořených uzlů (např. F+G) se ale zdají být vůči tomuto efektu imunní. Terminální větve jsou obarveny podle druhově-specifické rychlosti diverzifikace, vnitřní větve podle průměrné rychlosti diverzifikace dceřinných větví. Šedá mezikruží označují časové intervaly o délce 20 milionů let (takže např. pěvci jsou staří ~66 Ma, paleognáti ~64 Ma atd.). Vložený obrázek ukazuje rozmezí a frekvenci rychlostí diverzifikace napříč ptačími druhy. (Zdroj: Jetz et al. 2012: Figure 2)
3. Odhad fylogeneze: gory details
Zde právě přichází na řadu "kompletní" fylogeneze moderních ptáků. Autoři vytvořili fylogenetický strom čítající 9993 jednotlivých taxonů, a i když ne všechny zdroje by souhlasily, že je skutečně stoprocentně kompletní – Clements et al. [2012] např. odhadují počet ptačích druhů na 10 240 –, nemůže být sporu o tom, že rozsah díla je zcela bezprecedentní. Přirozeně se nabízí otázky, jak autoři dokázali "celoptačí" strom sestavit, když jsou genetická data k dispozici pouze od 6663 druhů (jak uvádějí sami autoři) – morfologie samozřejmě nic neřeší, protože nakódovat informativní znaky pro několik tisícovek taxonů by vyžadovalo strávit v muzejních depozitářích několik století – a když i daleko menší analýzy často staví současné počítače před velké problémy (Hackett et al. 2008). Sekce "Metody" v samotném textu studie postup objasňuje jen povrchně; to hlavní se nachází až v doplňkovém online materiálu. Tam autoři svou neradostnou situaci dále přibližují: software implementující sofistikované metody analýzy (např. BEAST, jednička v relaxovaných molekulárních hodinách, nebo MrBayes 3.2 a jeho působivé schopnosti v oblasti volby substitučních modelů) nezvládá odhadovat stromy s více než 1000 taxony, zatímco metody, které takto omezené nejsou (třeba PATHd8, vyhlazující tempo evoluce), trpí mnoha nedostatky: omezují tvar priorních rozdělení dob divergence okolo kalibračních fosilií a poskytují zavádějící odhady stáří uzlů. Autoři postupovali tak, že zkombinovali základní "kostru" (časově kalibrovaný strom velkoškálových příbuzenských vztahů v rámci ptáků) se samostatně odhadnutými fylogenezemi pro menší klady.
Topologie této kostry je založena na fylogenomických studiích ptáků jako celku (Ericson et al. 2006; Hackett et al. 2008) a pěvců (např. Barker et al. 2004; Jønsson et al. 2008). Zahrnuje celkem 158 koncových taxonů a 245 (u kostry odvozené z Hackett et al. 2008) nebo 249 (z Ericson et al. 2006) vnitřních uzlů, z nichž některé (ty s bootstrapem nad 70% nebo posteriorní pravděpodobnostní nad 0,95) si autoři vynutili po vlastní analýze (204 u Hackett et al., 212 u Ericson et al.). Autoři v tomto místě diskutují přetrvávající kontroverze ohledně dělení Neoaves na klady Metaves a Coronaves, které – jak je dobře známo – stojí a padá se zahrnutím 7. intronu β-fibrinogenu (Ericson et al. 2006; Hackett et al. 2008; Morgan-Richards et al. 2008). Signál podporující monofylii Metaves může být pouhým artefaktem špatně zarovnatelných dlouhých indelů, které jsou pro tento lokus typické (Morgan-Richards et al. 2008). Právě proto autoři důsledně používají dvě topologické kostry: ta, která je odvozená z Hackett et al. (2008), je založená na 19 lokusech a dichotomie Metaves/Coronaves na ní přítomná je; ta z Ericson et al. (2006: ESM-Figure 6) stojí na 4 genech (bez β-fibrinogenu) a Metaves ani Coronaves na ní nejsou. Topologie pěvců na obou podkladových stromech se řídí spíše jinými studiemi, hlavně pak Barker et al. (2004).
Ze 158 koncových taxonů 29 zahrnuje 4 a méně druhů, 129 naopak 5 a více (největší z nich dokonce plných 460). Každý z 9993 rozeznávaných druhů – autoři svůj výběr druhů odvozovali především ze světového seznamu Birdlife V3 z června 2010 – byl potom na základě předchozích fylogenetických studií zařazen do jednoho z těchto koncových taxonů. Následně Jetz et al. (2012) provedli fylogenetickou analýzu nad všemi druhy z jednoho koncového taxonu, pro které jsou dostupná genetická data, a celý postup opakovali, dokud nezískali fylogeneze pro všech 129 korunních kladů s více než 4 druhy. Při těchto analýzách si nevynucovali žádná omezení (constraints); data zde měla plnou svobodu diktovat výsledky. Každý z těchto 129 dílčích stromů byl také datován metodou relaxovaných molekulárních hodin.
Následně Jetz a spol. přistoupili k analýze 3323 taxonů, pro něž genetická data k dispozici nejsou. V tomto případě předpokládali, že taxonomie aproximuje fylogenezi: 2997 druhů bez dostupných DNA sekvencí je řazeno do "rodů", od nichž genetická data alespoň jeden druh máme. V takovém případě autoři prostě problematické taxony přilípli k jejich lépe prozkoumaným bratrancům. Tento postup má ale evidentní nevýhodu: předpokládá monofylii "rodů", přestože víme, že ptačí "rody" monofyletické velice často nejsou (Harshman 2007) a v případě pěvců mohou být dokonce druhy z jednoho "rodu" rozptýleny napříč několika "čeleděmi" (tamtéž). Jetz et al. (2012) proto sáhli po kompromisu: příslušenství k "rodu" ignorovali, pokud se ukázal být nonmonofyletickým na základě genetických dat, a dali druhu možnost zaujmout jakoukoli pozici uvnitř nejmenšího kladu obsahujícího všechny zástupce daného "rodu". Genetická data nejsou vůbec dostupná pro 214 "rodů" s celkem 326 druhy; tam se autoři spoléhali na zařazení do větších skupin. Logická otázka zní, jak vůbec do stromu zakomponovat taxony, pro něž nemáme žádná data (s výjimkou vágních priorů typu "X patří někam sem"). Zde se opět projevuje výhoda bayesovského přístupu: v případě absence sekvenčních dat Jetz et al. (2012) prostě nechali Markovovy řetězce Monte Carlo (MCMC) samplovat stromy z birth-death priorů (které jsou výhodné tím, že je lze kombinovat s "constraints" typu monofyletických "rodů"), přičemž tempo vymírání ("deaths" v birth-death modelu) bylo nastaveno na nulu. Není to proto, že by snad autoři věřili, že ptáci během své evoluci nevymírali, ale kvůli studiím ukazujícím, že homogenní tempo extinkce nezanechává žádný typický vzorec ve tvaru stromů.
V dalším kroku autoři zkombinovali stromy ze všech 129 kladů do jednoho celoptačího stromu, a to opět zvlášť pro taxony známé z genetických dat a zvlášť pro všechny taxony dohromady. První celoptačí strom zahrnoval 6670 taxonů (k 6663 ptákům, od nichž máme sekvence, se přidalo ještě 7 druhů, od nichž sice molekuly nemáme, ale které byly začleněny do výše zmíněných topologických koster, aby zajistily monofylii seriem, pokřovníkovitých, taxonů Mesitornis, Dasyornis a Daphoenositta, a aby pevně připoutaly ke stromu problematického strnada Převalského), ten druhý už kompletních 9993 taxonů. V tomto místě doplňkových dat mě docela pobavila poznámka o tom, jak topologická omezení pomohla omezit prostor možných topologií:
This excluded a portion of exceedingly unlikely possible topologies from our tree search. The remaining search space on topologies alone is >>10226, or effectively infinite.
Jetz et al. 2012: Supplementary Information p. 2
S celkovým 9993-taxonovým stromem už nám samozřejmě počet "constraints" podstatně narostl: k těm 245, resp. 249 páteřním se přidala ještě vynucená monofylie "rodů", nutná k zařazení druhů bez genetických dat; vynucená monofylie 57 "rodů", které obsahují víc než 1 druh, ale od žádného z nich nemáme sekvenční data; a vynucená monofylie "podčeledí", nutná k zařazení 214 "rodů" bez molekulárních dat. Celkem tedy počet neotestovaných předpokladů (či "priorů" nebo "constraints", jak jim říkají Jetz a spol.) narostl na 1770 uzlů v případě topologické kostry založené na Hackett et al. (2008) a 1774 v případě kostry postavené na Ericson et al. (2006) – tato čísla už šplhají k 18% celkového množství uzlů, což není málo. Znamená to vlastně, že jen nějakých 82% uzlů na výsledném 9993-taxonovém stromě je podloženo daty, která měli autoři v rukou. Jako velkého fandu bayesovských přístupů, zdůrazňujících užitečnost priorní informace pro jakýkoli odhad, mě to ale zas tak moc netrápí – vzhledem ke "cool factoru" studie jsem ochoten jí leccos odpustit. Navíc Jetz et al. (2012) ujišťují, že většina těchto omezení je velice slabá. Především zacházení s "rody" bez jakýchkoli genetických dat bylo ve studii velice liberální a řada z nich (103 z 214) dokonce mohla zaujmout jakoukoli pozici ve svém koncovém kladu.


Postup, předpoklady a "constraints" uplatněné při zahrnování taxonů bez genetických dat do stromu. (a) Pokud druh bez sekvenčních dat náleží do stejného "rodu" a, může se připojit do šedě vyznačené oblasti stromu. Druh "rodu" b, pro nějž sekvenční data také nemáme, se může vázat kamkoli do černé části stromu, protože b není monofyletický a černý podstrom je nejmenší skupina zahrnující všechny jeho zástupce známé z genetických dat. (b) Druhy bez genetických dat, patřící do rodů a, b, c, se připojí k druhům téhož "rodu", od nichž genetická data známe. Rod d ale není monofyletický, a tudíž se jeho druh bez genetických dat může vázat kamkoli v rámci kladu (a + b + c + d). Nesmí ale vstoupit do žádného dalšího rodu v rámci tohoto kladu, a proto se může připojovat jen na šedou část stromu. Předpokládejme dále, že chceme umístit druh patřící do "rodu", od nějž zcela chybí sekvenční data, a víme, že tento "rod" je příbuzný a a b. Pak se smí připojit pouze na větev X, protože do samotného kladu (a + b) už vstoupit nesmí. Kdybychom místo toho měli priorní informaci o příbuzenství s c, mohl by se naopak navázat pouze na větev Y a do samotného c už by vstoupit nesměl. (c) Opět předpokládejme, že umísťujeme druh patřící do "rodu" bez dostupných genetických dat. Víme, že je příbuzný s c a d. Potom smí vstoupit jen na šedě vyznačené větve. (Modifikováno z Jetz et al. 2012: Supplementary Methods Figure 2 a Supplementary Methods Figure 3)
V doplňkovém materiálu autoři také kriticky hodnotí, jak velkou část nejistoty o podobě ptačího rodokmenu se jejich studii podařilo obsáhnout ("obsáhnout nejistotu" je další bayesovský imperativ). Některé velkoškálové příbuzenské vztahy shrnuté v topologické kostře studie se jistě v budoucnu změní, a asi ani nebudeme muset moc dlouho čekat. McCormack et al. (2012), jejichž studie je stále ve fázi preprintu na arXivu, se s Hackett et al. (2008) i Ericsonem a spol. (2006) v několika bodech neshodnou. Změny tohoto typu by si také mohly vynutit přeřazení některých "rodů" do jiného ze 129 korunních kladů, které Jetz a spol. zavedli jako nenarušitelné jednotky s natvrdo nastaveným obsahem. Může se stát i to, že od druhů, které jsou v současnosti bez genetických dat, budou sekvence získány a ukáže se, že ve skutečnosti stojí mimo nejmenší klad vymezený již nyní osekvenovanými druhy z téhož rodu. Zcela nakonec může dojít i k tomu, že některé z "constraints" použitých k usazení "rodů" bez jakýchkoli genetických dat nebudou dostatečně liberální (např. kdyby se použité "podčeledě" ukázaly být polyfyletickými). Na všechny tyto situace dataset Jetze a spol. není připraven. Všechnu další nejistotu o ptačí fylogenezi však již pojmout dokáže.
K získání 129 dílčích stromů autoři použili BEAST, který bayesovskými metodami simultánně odhaduje fylogenezi a časově kalibrované délky větví a umožňuje důkladně si pohrát s priorními rozděleními pro data divergencí (viz nedávný článek na blogu). Jako outgroup pro analýzu každého ze 129 menších kladů byl vždy použit jeho nejbližší a druhý nejbližší příbuzný na topologické kostře. Konkrétní druhy pak byly z těchto kladů vybrány tak, aby od nich bylo k dispozici co nejvíce genů, které jsou nejčastěji osekvenovány pro taxony z ingroup. Vzorek stromů získaný z MCMC autoři většinou shrnovali jako 95% konsenzus (jen klady přítomné na 95% a více stromech ze vzorku se dostanou na finální strom), ale v sedmi případech zvolili 75% a v jednom (u tyranovitých a příbuzných) dokonce jen 50% konsenzus, aby se zvýšilo rozlišení a taxony nezastoupené genetickými daty mohly být přesněji usazeny. Pokud se na těchto stromech vyskytly polytomie, autoři se je snažili rozřešit tak, aby zachovali monofylii "rodů" a aby skupiny, které vymezovaly postavení "rodů" nezastoupených molekulárními daty, měly co nejmenší rozsah.
Aby získali kompletní topologickou kostru, na kterou by šlo "rozvěsit" 129 dílčích stormů, museli Jetz et al. (2012) něco udělat s těmi několika desítkami uzlů, které Ericson et al. (2006) ani Hackett et al. (2008) nevykryli s podporou dost vysokou na to, aby bylo rozumné použít je jako tvrdé "constraints". Řešením je vlastní fylogenetická analýza: autoři 15 genů, zahrnujících široký výběr jaderných intronů, exonů i mitochondriálních protein-kódujících genů. Z každého ze 129 větších kladů byl vybrán jeden reprezentativní druh a samplováno rovněž bylo 21 kladů obsahujících jeden až čtyři druhy. Jak už bylo uvedeno výše, od 7 druhů nebyla genetická data dostupná, přesto ale byly natvrdo dosazeny do topologické kostry. Samplování taxonů bylo samozřejmě opět nastaveno tak, aby od každého druhu bylo k dispozici co možná nejvíc genů. K alignmentu byla použita pokročilá metoda PRANK (krátce zmíněná na blogu zde), dataset byl rozdělen do 13 oddílů s odlišnými modely evoluce. Celkově datová matice dosáhla délky 13 277 párů bází. Rizikem ovšem zůstává nedostatečná rozhodnost dat: když se sekvence od jednotlivých taxonů budou překrývat jen velmi málo, může se stát, že jejich přesun napříč stromem neovlivní věrohodnost, což by představovalo velký problém (Sanderson et al. 2010). Rozhodnost měřená jako podíl větví, které analýza dokázala rozlišit na 1000 náhodně vybraných topologií, ale byla vysoká dostatečně (> 98%).
K molekulárnímu datování autoři využívají 13 fosilií, mezi nimiž najdeme už klasické a všeobecně oblíbené kalibrační body typu vegavise (z kmenové linie kachnovitých) a waimanua (z kmenové linie tučňáků). Do analýzy je autoři zakomponovali pomocí lognormálních priorních rozdělení s tvrdým minimem na věku fosilie a měkkým maximem na stáří spodnokřídového bazálního ornituromorfa (O'Connor et al. 2011) Gansus yumenensis. (Tato horní hranice je mimochodem skutečně pro všechny divergence stejná: stáří 110 milionů let může dosáhnout jakákoli skupina, od tučňáků přes rorýse a dudky až po papouščí klad Psittacoidea, budou-li si to data žádat.) Lognormální rozdělení je podle Jetze a spol. zvlášť vhodné proto, že disponuje tvrdou spodní hranicí (takže divergence nemohou být mladší než fosilní kalibrační body) a střední hodnota (na níž je pravděpodobnost divergence nejvyšší) se nachází kousek za tímto minimem, což koresponduje s faktem, že divergence musejí být nutně starší než fosilie, které jsou jejich produkty. Stáří kořene stromu, tzn. divergence mezi paleognáty a neognáty, bylo rovněž modelováno pomocí lognormálního rozdělení s měkkým minimem na 66 Mya (stáří vegavise) a měkkým maximem na 110 Mya (5% celkové hustoty pravděpodobnosti za maximem); ostatní divergence měly vlastní minima a za maximem se nacházelo 2,5% celkové hustoty.
4. Modely, rychlosti a nečekané adaptivní radiace
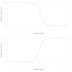
Podíváme-li se na výsledný datovaný strom 9993 druhů a zaměříme se na to, kolik linií existovalo v kterémkoli daném čase, zjistíme, že celková diverzifikace (speciace minus extinkce) u moderních ptáků začala narůstat před 50 miliony let a ustala nejdříve před 5 miliony let. Jde o dobrý příklad situace, kdy je globální vzorec odlišný od vzorců lokálních: studie menších kladů (Phillimore & Price 2008; Rabosky & Lovette 2008a) ukazují, že diverzifikace se časem snižuje, což souvisí s výše zmíněným hustotně-závislým modelem. V takových případech se zřejmě skupině na počátku dostane možnosti zaplnit volné niky, o které pak jednotlivé linie dlouhodobě kompetují.

K volbě mezi jednotlivými modely diverzifikace Jetz a spol. využili Akaikeho informační kritérium (AIC). A skutečně je z čeho vybírat. Jak ukazuje např. výborný, volně dostupný text paleontologa Davida Bapsta, modelování tempa diverzifikace z molekulárních fylogenezí je extrémně rychle se rozvíjejícím oborem a modelů přibývá obrovským tempem. Autoři současné studie jich zkoušejí 9. "Pure birth" je z nich nejjednodušší a obsahuje jediný parametr, udávající rychlost speciace (modelované jako Poissonův proces); vývojové linie podle něj nikdy nevymírají. "Birth-death" model už alespoň započítává tempo vymírání coby druhý parametr, stále ale předpokládá, že rychlosti speaciace a extinkce jsou konstantní. Hustotně-závislý logistický model se parametru rychlosti vymírání opět zbavuje a místo něj zavádí do výpočtu tempa diverzifikace počet linií na časovém horizontu našeho zájmu a pomocný parametr K (Rabosky 2006; Rabosky & Lovette 2008a). Exponenciální hustotně-závislý model vypouští K a zavádí x, upravující velikost změn speciační rychlosti jako funkci počtu linií. SPVAR model opět počítá jen se speciací a vymírání, ale dovoluje prvnímu procesu měnit rychlost, čímž dospějeme ke třem parametrům – blízce příbuzný je model EXVAR, který ponechává tempo speciace konstantní a změny povoluje jen v tempu extinkce, čímž opět dospívá ke třem parametrům (Rabosky & Lovette 2008b). Model TreePar patří mezi "birth-death-shift" modely: nedovoluje rychlostem speciace a vymírání měnit se ve spojitém čase, jako je tomu ve SPVARu a EXVARu, ale umožňuje mezi různými rychlostmi diskrétní přechody. Autoři vyzkoušeli dvě varianty: s jedním a se dvěma přechody v rámci celých ptáků (víc jich AIC nepovolilo). Konečně MEDUSA funguje velice podobně, ale snaží se nějak vhodně umístit 20 diskrétních změn rychlosti diverzifikace na různé větve.
Asi nepřekvapí, že na evoluci kladu s tak bohatou evoluční historií, jakým jsou moderní ptáci, nejlépe sedí ten nejkomplikovanější model: MEDUSA s 20 diskrétními posuny v tempu speciace. Vždy se přitom jednalo o zrychlení, a ve většině případů k nim došlo v posledních ~50 milionech let. Některé z těchto posunů byly optimizovány na bázi vysoce diverzifikovaných kladů, jejichž rozmanitost už byla dříve dána do souvislosti s klíčovými morfologickými a etologickými apomorfiemi nebo s příležitostí obsadit nová prostředí. Sem patří třeba zrychlená diverzifikace kolibříků, papoušků a četných linií pěvců. Další zrychlení tempa speciace byla identifikována na bází mladých skupin, procházejících v současnosti rapidní radiací (0,25 nových druhů za 1 milion let a rychlejší) – u hrnčiříkovitých (Furnariidae) a výše zmíněných anomálních kruhoočkovitých (Zosteropidae) to nepřekvapí, ale rapidní radiace v rámci kachnovitých (stáří uzlu 10,8 Ma; tempo speciace 0,28 druhů/milion let) nebo racků (4,6 Ma; 0,74 druhů/milion let) dosud rozeznány nebyly. Už zde se evidentně začínají projevovat výhody kompletní datované ptačí fylogeneze. Jetz et al. (2012) ukazují, že tyto nedávno radiující skupiny mají mezi 44 a 355 druhy, a – zajímavěji – neomezují se na žádnou konkrétní část fylogenetického stromu ani pevninu; najdeme je od galloanseranů po sylvioidní pěvce. Právě tyto "výbuchy diverzity" rozptýlené napříč ptačím stromem způsobují, že rychlost diverzifikace ptáků jako celku roste. Zároveň je důležité podotknout, že krom zmiňovaných 20 diskrétních posunů je rychlost diverzifikace považována za konstantní. Její nárůsty a opětovné poklesy v menších kladech, jaké zdokumentovali Rabosky & Lovette (2008a) či Phillimore & Price (2008), jsou tedy s modelem předkládaným Jetzem a spol. dobře kompatibilní, protože konstantní tempo lze interpretovat i jako vzájemné vyrušení lokálních zrychlení a zpomalení.
5. Geografie: na ostrovním výskytu záleží, na zeměpisné šířce ne
Aby lépe pochopili důsledky posunů v rychlosti diverzifikace na dnešní geografické odchylky v ptačí diverzitě, zavedli Jetz et al. (2012) nové měřítko: druhově specifickou rychlosti diverzifikace (species-level lineage diversification rate, dále jen DR). Jde o upravenou verzi měřítka známého jako "Equal Splitting" (ES; Redding & Mooers 2006), které – jak název napovídá – rozděluje délku rodičovské větve rovnoměrně mezi obě větve dceřinné. Celkově tak umožňuje stručně shrnout evoluční historii druhu: hodnota ES je součet délek větví od kořene stromu ke špičce (větvi samotného druhu), přičemž každou další větev násobíme 0,5. DR je pak prostě 1/ES. Jak by mělo být z popisu výše patrné, samotná možnost toto měřítko aplikovat je jedním z dalších kouzel, které umožňuje kompletně samplovaná fylogeneze. Hodnoty DR pro jednotlivé druhy se pohybují mezi 0,01 a 4,66 druhu na milion let a přibližně odpovídají lognormálnímu rozdělení. Toto rozdělení ukazuje velké rozdíly v rychlostech diverzifikace mimo 20 hlavních posunů a umožňuje identifikovat stále "aktivní" místa v nedávno radiujících skupinách.
Hlavní geografické rozdíly související s tempem diverzifikace jsou překvapivě mezi západní a východní polokoulí a nesouvisejí se zeměpisnou šířkou. Na východní polokouli (na východ od 28° západní délky) je DR podstatně nižší než na té západní (0,145 oproti 0,177), zatímco jižní a severní polokoule vykazují průměrné tempo diverzifikace téměř shodné (0,154 a 0,161, v tomto pořadí). Avifauny Afriky, Madagaskaru, jihovýchodní Asie a Austrálie vykazují nižší DR, než je globální průměr (pod 0,12 oproti 0,16), a 25% druhů s nejvyššími rychlostmi diverzifikace se na jejich složení podílí méně, než bychom očekávali (z méně než 15%). Tento překvapivý vzorec by opět mohla vysvětlit hustotně závislá diverzifikace: obsazení dostupného ekologického prostoru dávnými radiacemi a následná kompetice, doprovázená klesajícím tempem diverzifikace (což navrhl např. Ericson 2012). Přesně opačně na tom jsou severní části Severní Ameriky, jihozápadní Jižní Amerika a některé oblasti severní Asie, kde najdeme vysoká DR a relativně vysoká zastoupení ptáků s nejvyššími rychlostmi diverzifikace. Právě v těchto regionech najdeme rapidní radiace zmiňované výše, jako ty u kachnovitých, racků, datlů a některých linií pěvců, a od pliocénu (5.3 Mya) zde kolísá klima, což doprovází střídání mokřadů a lesů. DR hodnoty ale můžeme uchopit i za opačný konec a místo hledání vysvětlení pro pozorované rozdíly je použít k testování zajímavých hypotéz – např. té, podle níž ostrovy a podobné izolované lokality usnadňují rapidní radiace, neboť znesnadňují genový tok a poskytují kolonizátorům nový ekologický prostor k zaplnění. Data to skutečně potvrzují: vyloučíme-li ptáky žijící na otevřeném moři, zjistíme, že taxony, jejichž oblast výskytu se více než z půlky rozkládá na ostrovech, mají průměrné DR daleko vyšší než ptáci převážně pevninští (0,203 oproti 0,153). Řada ostrovů vykazuje neobvykle vysoké rychlosti diverzifikace – především to platí pro pěvce v Karibiku, severním Atlantiku a Pacifiku (např. opět kruhoočkovití).

Fylogeneticky vzato se ukazuje, že pěvci mají vyšší průměrnou rychlost diverzifikace než ostatní ptáci (0,179 oproti 0,131), což by, jak autoři podotýkají, nemělo překvapovat už jen proto, že jejich prostě víc (z celkových 9993 druhů je 5966 pěvců). Toto zjištění odpovídá i skutečnosti, že jeden z posunů v tempu diverzifikace MEDUSA lokalizovala právě na bázi pěvců, konkrétně uzel Eupasseres (sensu Mayr 2005, 2011: zpěvní [Oscines] + křikaví [Suboscines] neboli všichni pěvci mimo pokřovníkovitých [Acanthisittidae]). Vysoké rychlosti diverzifikace pozorované v některých částech světa tedy táhnou především pěvci, ačkoli je pravda, že pěvci i ostatní ptáci pokrývají široké spektrum diverzifikačních rychlostí a obě skupiny také vykazují zvýšené tempo na ostrovech. Jetz et al. (2012) přesto odhalují zajímavý efekt: vyšší rychlosti diverzifikace v Asii a Severní Americe táhnou primárně nepěvčí ptáci (racci, datli, Aequornithes), zatímco na vysokém průměrném tempu diverzifikace v Jižní Americe a především pak jižní části hlavního pásma deštných lesů se podílejí prakticky výhradně radiace pěvců, hlavně hrnčiříků a klouzálků (vytvářejících klad označovaný buď jako Furnariidae sensu lato, nebo štěpený do "čeledí" Dendrocolaptidae, Scleruridae a Furnariidae sensu Moyle et al. 2009b). Autoři tento vzorec vysvětlují zvýšením plochy oblastí s mírným podnebím v posledních 10 až 20 milionech let a v nedávné době i ústupem zalednění ve vyšších zeměpisných šířkách.
Tato zjištění kontrastují s nedávnými studiemi, podle níž rychlosti speciace i vymírání výrazně vzrůstají se zeměpisnou šířkou (Weir & Schluter 2007). Přestože většina druhů v horním kvartilu rychlosti diverzifikace má skutečně střední zeměpisnou šířku svého areálu mezi 30° severní a 30° jižní šířky, totéž platí pro všechny druhy (tropy jsou prostě co do počtu taxonů bohatší) a v nížinatých tropech je co doo počtu překonávají pomalu diverzifikující druhy. Průměrná rychlosti diverzifikace nevykazuje ve vztahu k zeměpisné šířce žádný výrazný trend a není signifikantně odlišná ani pro intra- verzus extra-tropické střední zeměpisné šířky. Rozdíly v diverzitě na různých zeměpisných šířkách tak nelze vysvětlit prostými rozdíly v celkové rychlosti diverzifikace (tedy že by v tropech měla speciace "náskok" před extinkcí), contra Weir & Schluter (2007). Potvrzena se tím naopak zdá být hypotéza, že větší diverzita v tropech je způsobena delší dobou existence a větší rozlohou deštných lesů oproti jiným prostředím (Jetz & Fine 2012) – jak připomíná Ricklefs (2012) v krátkém shrnutí a diskuzi nové práce, ještě 30 milionů let zpátky byla Země převážně tropická, než se začaly rozšiřovat oblasti mírného a boreálního klimatu. Zatímco na rovníku samotném je v průměrné rychlosti diverzifikace patrný propad, mezi 10° a 15° zeměpisné šířky hodnoty narůstají (což Jetz a spol. opět dávají do souvislosti s vyšší klimatickou dynamikou v posledních 10 milionech let) a ve vyšších šířkách opět klesají. Autoři ale upozorňují, že tato šířková variace je méně významná než variace mezi polokoulemi, mezi pevninskými a ostrovními druhy a konečně i s výskytem, rozšířením a evoluční dynamikou konkrétních ptačích kladů v tom kterém místě. Ricklefs (2012) připomíná, že absence zjevných rozdílů v tempu diverzifikace v závislosti na zeměpisné šířce byla nedávno rovněž doložena u savců (Soria-Carrasco & Castresana 2012).

6. Robustnost a nejistota
Důležité je i to, že všechny tyto výsledky jsou robustní vůči volbě topologické kostry (Ericson et al. 2006: ESM-Figure 6 vs. Hackett et al. 2008) a vůči zahrnutí druhů s nedostatkem dat. Jetz et al. (2012) v tomto místě také konstatují, že jejich strom, snažící se najít vyvážený kompromis mezi obsáhnutím zbývající nejistoty ohledně ptačí fylogeneze a integrací nejnovějších taxonomických dat, nebude použitelný pro všechny účely. Absolutní rychlosti diverzifikace se zase odvíjejí od stáří korunního kladu ptáků (Neornithes) a to zas přímo závisí na použití konkrétních kalibračních fosilií. (Odhad otištěný ve studii jej mimochodem stanovuje na ~108 Ma, což je příjemné a rozhodně realističtější než 127–131 Ma, které nedávno nahlásili Haddrath & Baker [2012].) Především v tropech se pak jistě skrývá ještě mnoho kryptických druhů, které mohou dále změnit průměrné rychlosti diverzifikace oblasti. Nakonec Jetz et al. (2012) přiznávají i omezení vyplývající z toho, že jejich studie zahrnuje jen žijící taxony a nemůže tak obsáhnout informace o kompletně vymřelých kladech typu novozélandských moa.
7. Ptačí accelerando: anomálie, nebo zákonitost skrytá pod neúplným samplingem?
Geografické rozdíly, které z dat vyvstaly teprve s první globální analýzou rozdílů v druhově-specifických rychlostech diverzifikace v prostoru i čase, se zdají být velice výrazné a naznačují, že ptáci jsou v nějakém ohledu výjimeční: jejich rostoucí tempo diverzifikace v posledních 50 milionech let je velmi zvláštní. Jetz et al. (2012) spekulují, že adaptivní "nad-zóna", do níž ptáci nejpozději od raného eocénu diverzifikují, ještě pořád nemusí být nasycena a příležitostí pro další expanzi v ní přibývá. Konzervativnější vysvětlení je, že nasycení obsazeného ekologického prostoru, indikované analýzami jiných skupin, je jen artefaktem nedostatečného samplingu. Saturace je ale pravděpodobně stále reálným fenoménem u menších kladů s omezeným geografickým a/nebo ekologickým rozsahem (Phillimore & Price 2008; Rabosky & Lovette 2008a). Jetz a spol. navrhují tyto problémy řešit porovnáváním kompletních globálních stromů, jako je ten jejich, s lokálními analýzami menších skupin. Předpovídají, že ačkoli na menších geografických a ekologických meřítkách rychlost diverzifikace skutečně s časem klesat v souladu s hustotně-závislým modelem, tyto menší vzorce se nesloží a nevyprodukují stejný vzorec na větším, globálním měřítku. Co platí jednotlivě pro budníčky, lesňáčkovité a trogony neplatí pro skupinu z nich složenou; jednotlivé adaptivní radiace mohou být ve svém rozsahu omezeny, ale na diverzitu skupiny, která je produkuje, se žádný takový limit nevztahuje. Ptačí biodiverzita je víc než součet jejích částí.
8. Pagel (2012): první kritika
Současně se studií byl v Nature otištěno ještě citované shrnutí z pera Roberta Ricklefse, který má sám na kontě působivé množství studií o vzorcích ptačí (a především pak pěvčí) biodvierzity: ukázal, které pěvčí klady jsou rozmanitější nebo méně rozmanité, než bychom očekávali, a ukázal, že anomálie mají původ spíš v biogeografii než klíčových inovacích (Ricklefs 2003); doložil korelaci mezi morfologickou variabilitou a počtem druhů, naznačující vliv kladogeneze na evoluci znaků (Ricklefs 2004), a ukázal, že druhově chudé klady lze většinou nalézt na periferii pěvčího morfoprostotu (Ricklefs 2005). Za druhým textem reagujícím na megafylogenezi Jetze a spol. stojí fylogenetik Mark Pagel. Ten zdůrazňuje, že studie typu té od Jetze a spol. jsou nesmírně cenné v tom, že umožňují rekonstruovat řadu dalších aspektů evoluce než je "holá" fylogeneze, včetně ancestrálních znakových stavů, tempa speciace nebo rychlosti morfologických změn. Vedou tedy k úplnějšímu pochopení evolučních událostí a procesů tvarujících historii skupiny, a obrovská statistická síla vyplývající ze samplingu, který je hustý i široký současně, umožňuje zamítat hypotézy s větším stupněm jistoty.
Poté ale Pagel (2012) přichází ke kritice. Upozorňuje, že spoléhání se na taxonomii při umísťování druhů nezastoupených genetickými daty a na předchozí fylogenetické studie při konstrukci topologické kostry neumožnilo dostatečně prozkoumat "the possible universe of avian relatedness" (Pagel 2012:2). Místo toho by druhy, od nichž máme molekulární data, měly mít možnost shlukovat se tak, jak diktují data (bez předem vynucených uzlů), a druhy, od nichž sekvence nemáme, by měla do stromu zařadit fylogenetická analýza jiných typů dat (morfologie, chování). Osobně si myslím, že ani jedna z těchto výtek není zvlášť užitečná; jistě je to pravda, ale nasbírat dostatečné množství non-molekulárních dat pro fylogenetickou analýzu 3330 ptáků se nezdá být příliš reálné. Stejně tak výhrada, že Jetz et al. (2012) do své studie nezahrnuli fylogenetická data z nedávné studie o zrychlené speciaci u barevně polymorfických ptáků (Hugall & Stuart-Fox 2012), která přišla s konvenční fylogenetickou analýzou zhruba 4000 druhů pěvců, je spíš jen formální. Vzhledem k tomu, že práce Hugalla a Stuart-Fox vyšla online 9. května 2012 a studie Jetze a spol. 31. října 2012, je těžké vidět, jak by ta druhá mohla zahrnout jakákoli data z té první.
Jedinou skutečně zajímavou kritiku představuje ta nejvěcnější. Většina ze závěrů Jetze a spol. je založena na jejich vlastním měřítku druhově specifického diverzifikačního tempa (DR), které je pouze převrácenou hodnotou měřírka ES, sčítajícího zlomky délek větví vedoucích od daného druhu ke kořenu stromu. Důležité je tedy spolehlivě odhadnout délky větví, a právě tam je u metod využívajících relaxovaných molekulárních hodin (jako je BEAST) potřeba velmi velká opatrnost (Venditti et al. 2010). Pagel navíc připomíná, že v tomto případě je ohromný rozsah stromu Jetze a spol. spíš dalším důvodem k obavám než výhodou: i malá chyba v odhadu délek větví může mít na tak velké fylogenezi velký dopad na konečné DR, a trendy odvozené z těchto hodnot pak nemusejí být vůbec reálné. Doporučuje, aby se souvislosti mezi klimatem, ekologií a diverzifikačními rychlostmi, které Jetz et al. (2012) předkládají, testovaly standardní komparativní metodou, tzn. hledáním dvojic nezávislých znaků, které se vyvíjejí pospolu, a na samotnou studii pohlíží spíš jako na první krůček na cestě k úplné fylogenezi ptáků a porozumění ptačí biodiverzitě.
9. V médiích
First ever family tree for all living birds reveals evolution and diversification (Science Daily) – podotýká, že globální fylogeneze se může hodit i pro stanovování priorit v konzervační biologii.
Exhaustive family tree for birds shows recent, rapid diversification (Science Daily)
Are birds accelerating their rate of evolution? (Science 2.0)
10. Acknowledgments
Děkuji Rafaelu Maiovi za zaslání kompletního PDF nové studie.
Zdroje:
Aby získali kompletní topologickou kostru, na kterou by šlo "rozvěsit" 129 dílčích stormů, museli Jetz et al. (2012) něco udělat s těmi několika desítkami uzlů, které Ericson et al. (2006) ani Hackett et al. (2008) nevykryli s podporou dost vysokou na to, aby bylo rozumné použít je jako tvrdé "constraints". Řešením je vlastní fylogenetická analýza: autoři 15 genů, zahrnujících široký výběr jaderných intronů, exonů i mitochondriálních protein-kódujících genů. Z každého ze 129 větších kladů byl vybrán jeden reprezentativní druh a samplováno rovněž bylo 21 kladů obsahujících jeden až čtyři druhy. Jak už bylo uvedeno výše, od 7 druhů nebyla genetická data dostupná, přesto ale byly natvrdo dosazeny do topologické kostry. Samplování taxonů bylo samozřejmě opět nastaveno tak, aby od každého druhu bylo k dispozici co možná nejvíc genů. K alignmentu byla použita pokročilá metoda PRANK (krátce zmíněná na blogu zde), dataset byl rozdělen do 13 oddílů s odlišnými modely evoluce. Celkově datová matice dosáhla délky 13 277 párů bází. Rizikem ovšem zůstává nedostatečná rozhodnost dat: když se sekvence od jednotlivých taxonů budou překrývat jen velmi málo, může se stát, že jejich přesun napříč stromem neovlivní věrohodnost, což by představovalo velký problém (Sanderson et al. 2010). Rozhodnost měřená jako podíl větví, které analýza dokázala rozlišit na 1000 náhodně vybraných topologií, ale byla vysoká dostatečně (> 98%).
K molekulárnímu datování autoři využívají 13 fosilií, mezi nimiž najdeme už klasické a všeobecně oblíbené kalibrační body typu vegavise (z kmenové linie kachnovitých) a waimanua (z kmenové linie tučňáků). Do analýzy je autoři zakomponovali pomocí lognormálních priorních rozdělení s tvrdým minimem na věku fosilie a měkkým maximem na stáří spodnokřídového bazálního ornituromorfa (O'Connor et al. 2011) Gansus yumenensis. (Tato horní hranice je mimochodem skutečně pro všechny divergence stejná: stáří 110 milionů let může dosáhnout jakákoli skupina, od tučňáků přes rorýse a dudky až po papouščí klad Psittacoidea, budou-li si to data žádat.) Lognormální rozdělení je podle Jetze a spol. zvlášť vhodné proto, že disponuje tvrdou spodní hranicí (takže divergence nemohou být mladší než fosilní kalibrační body) a střední hodnota (na níž je pravděpodobnost divergence nejvyšší) se nachází kousek za tímto minimem, což koresponduje s faktem, že divergence musejí být nutně starší než fosilie, které jsou jejich produkty. Stáří kořene stromu, tzn. divergence mezi paleognáty a neognáty, bylo rovněž modelováno pomocí lognormálního rozdělení s měkkým minimem na 66 Mya (stáří vegavise) a měkkým maximem na 110 Mya (5% celkové hustoty pravděpodobnosti za maximem); ostatní divergence měly vlastní minima a za maximem se nacházelo 2,5% celkové hustoty.
4. Modely, rychlosti a nečekané adaptivní radiace
Podíváme-li se na výsledný datovaný strom 9993 druhů a zaměříme se na to, kolik linií existovalo v kterémkoli daném čase, zjistíme, že celková diverzifikace (speciace minus extinkce) u moderních ptáků začala narůstat před 50 miliony let a ustala nejdříve před 5 miliony let. Jde o dobrý příklad situace, kdy je globální vzorec odlišný od vzorců lokálních: studie menších kladů (Phillimore & Price 2008; Rabosky & Lovette 2008a) ukazují, že diverzifikace se časem snižuje, což souvisí s výše zmíněným hustotně-závislým modelem. V takových případech se zřejmě skupině na počátku dostane možnosti zaplnit volné niky, o které pak jednotlivé linie dlouhodobě kompetují.

Odhady (a) rychlosti celkové diverzifikace (speciace minus extinkce) a (b) rychlosti speciace pro moderní ptáky jako celek, počítané v počtu druhů za milion let po 5 milionů let dlouhých intervalech ("zuby" v grafech). Šedá barva vyznačuje prostor mezi 5% a 95% kvantilem pro 525 otestovaných stromů, průměr je vyznačen černou čarou. Období starší než 67,5 Ma nebo a mladší než 2,5 Ma nejsou pro nedostatek dat v grafech zobrazena: v prvním případě počet samostatných linií klesl pod 30 a dataset už nemá dostatečnou statistickou sílu, v druhém případě je obtížné vyhnout se problémům s právě probíhajícími speciačními událostmi. (c) Graf zobrazující počet linií koexistujících na daném časovém horizontu – opět je patrné, že rychlost diverzifikace ptáků se přes posledních 50 milionů let neustále zvyšuje (barevné značení stejné jako v předchozích grafech). Na pozadí výše přetištěný fylogenetický strom. (Zdroj: Jetz et al. 2012: Figure 1)
K volbě mezi jednotlivými modely diverzifikace Jetz a spol. využili Akaikeho informační kritérium (AIC). A skutečně je z čeho vybírat. Jak ukazuje např. výborný, volně dostupný text paleontologa Davida Bapsta, modelování tempa diverzifikace z molekulárních fylogenezí je extrémně rychle se rozvíjejícím oborem a modelů přibývá obrovským tempem. Autoři současné studie jich zkoušejí 9. "Pure birth" je z nich nejjednodušší a obsahuje jediný parametr, udávající rychlost speciace (modelované jako Poissonův proces); vývojové linie podle něj nikdy nevymírají. "Birth-death" model už alespoň započítává tempo vymírání coby druhý parametr, stále ale předpokládá, že rychlosti speaciace a extinkce jsou konstantní. Hustotně-závislý logistický model se parametru rychlosti vymírání opět zbavuje a místo něj zavádí do výpočtu tempa diverzifikace počet linií na časovém horizontu našeho zájmu a pomocný parametr K (Rabosky 2006; Rabosky & Lovette 2008a). Exponenciální hustotně-závislý model vypouští K a zavádí x, upravující velikost změn speciační rychlosti jako funkci počtu linií. SPVAR model opět počítá jen se speciací a vymírání, ale dovoluje prvnímu procesu měnit rychlost, čímž dospějeme ke třem parametrům – blízce příbuzný je model EXVAR, který ponechává tempo speciace konstantní a změny povoluje jen v tempu extinkce, čímž opět dospívá ke třem parametrům (Rabosky & Lovette 2008b). Model TreePar patří mezi "birth-death-shift" modely: nedovoluje rychlostem speciace a vymírání měnit se ve spojitém čase, jako je tomu ve SPVARu a EXVARu, ale umožňuje mezi různými rychlostmi diskrétní přechody. Autoři vyzkoušeli dvě varianty: s jedním a se dvěma přechody v rámci celých ptáků (víc jich AIC nepovolilo). Konečně MEDUSA funguje velice podobně, ale snaží se nějak vhodně umístit 20 diskrétních změn rychlosti diverzifikace na různé větve.
Asi nepřekvapí, že na evoluci kladu s tak bohatou evoluční historií, jakým jsou moderní ptáci, nejlépe sedí ten nejkomplikovanější model: MEDUSA s 20 diskrétními posuny v tempu speciace. Vždy se přitom jednalo o zrychlení, a ve většině případů k nim došlo v posledních ~50 milionech let. Některé z těchto posunů byly optimizovány na bázi vysoce diverzifikovaných kladů, jejichž rozmanitost už byla dříve dána do souvislosti s klíčovými morfologickými a etologickými apomorfiemi nebo s příležitostí obsadit nová prostředí. Sem patří třeba zrychlená diverzifikace kolibříků, papoušků a četných linií pěvců. Další zrychlení tempa speciace byla identifikována na bází mladých skupin, procházejících v současnosti rapidní radiací (0,25 nových druhů za 1 milion let a rychlejší) – u hrnčiříkovitých (Furnariidae) a výše zmíněných anomálních kruhoočkovitých (Zosteropidae) to nepřekvapí, ale rapidní radiace v rámci kachnovitých (stáří uzlu 10,8 Ma; tempo speciace 0,28 druhů/milion let) nebo racků (4,6 Ma; 0,74 druhů/milion let) dosud rozeznány nebyly. Už zde se evidentně začínají projevovat výhody kompletní datované ptačí fylogeneze. Jetz et al. (2012) ukazují, že tyto nedávno radiující skupiny mají mezi 44 a 355 druhy, a – zajímavěji – neomezují se na žádnou konkrétní část fylogenetického stromu ani pevninu; najdeme je od galloanseranů po sylvioidní pěvce. Právě tyto "výbuchy diverzity" rozptýlené napříč ptačím stromem způsobují, že rychlost diverzifikace ptáků jako celku roste. Zároveň je důležité podotknout, že krom zmiňovaných 20 diskrétních posunů je rychlost diverzifikace považována za konstantní. Její nárůsty a opětovné poklesy v menších kladech, jaké zdokumentovali Rabosky & Lovette (2008a) či Phillimore & Price (2008), jsou tedy s modelem předkládaným Jetzem a spol. dobře kompatibilní, protože konstantní tempo lze interpretovat i jako vzájemné vyrušení lokálních zrychlení a zpomalení.
5. Geografie: na ostrovním výskytu záleží, na zeměpisné šířce ne
Aby lépe pochopili důsledky posunů v rychlosti diverzifikace na dnešní geografické odchylky v ptačí diverzitě, zavedli Jetz et al. (2012) nové měřítko: druhově specifickou rychlosti diverzifikace (species-level lineage diversification rate, dále jen DR). Jde o upravenou verzi měřítka známého jako "Equal Splitting" (ES; Redding & Mooers 2006), které – jak název napovídá – rozděluje délku rodičovské větve rovnoměrně mezi obě větve dceřinné. Celkově tak umožňuje stručně shrnout evoluční historii druhu: hodnota ES je součet délek větví od kořene stromu ke špičce (větvi samotného druhu), přičemž každou další větev násobíme 0,5. DR je pak prostě 1/ES. Jak by mělo být z popisu výše patrné, samotná možnost toto měřítko aplikovat je jedním z dalších kouzel, které umožňuje kompletně samplovaná fylogeneze. Hodnoty DR pro jednotlivé druhy se pohybují mezi 0,01 a 4,66 druhu na milion let a přibližně odpovídají lognormálnímu rozdělení. Toto rozdělení ukazuje velké rozdíly v rychlostech diverzifikace mimo 20 hlavních posunů a umožňuje identifikovat stále "aktivní" místa v nedávno radiujících skupinách.
Hlavní geografické rozdíly související s tempem diverzifikace jsou překvapivě mezi západní a východní polokoulí a nesouvisejí se zeměpisnou šířkou. Na východní polokouli (na východ od 28° západní délky) je DR podstatně nižší než na té západní (0,145 oproti 0,177), zatímco jižní a severní polokoule vykazují průměrné tempo diverzifikace téměř shodné (0,154 a 0,161, v tomto pořadí). Avifauny Afriky, Madagaskaru, jihovýchodní Asie a Austrálie vykazují nižší DR, než je globální průměr (pod 0,12 oproti 0,16), a 25% druhů s nejvyššími rychlostmi diverzifikace se na jejich složení podílí méně, než bychom očekávali (z méně než 15%). Tento překvapivý vzorec by opět mohla vysvětlit hustotně závislá diverzifikace: obsazení dostupného ekologického prostoru dávnými radiacemi a následná kompetice, doprovázená klesajícím tempem diverzifikace (což navrhl např. Ericson 2012). Přesně opačně na tom jsou severní části Severní Ameriky, jihozápadní Jižní Amerika a některé oblasti severní Asie, kde najdeme vysoká DR a relativně vysoká zastoupení ptáků s nejvyššími rychlostmi diverzifikace. Právě v těchto regionech najdeme rapidní radiace zmiňované výše, jako ty u kachnovitých, racků, datlů a některých linií pěvců, a od pliocénu (5.3 Mya) zde kolísá klima, což doprovází střídání mokřadů a lesů. DR hodnoty ale můžeme uchopit i za opačný konec a místo hledání vysvětlení pro pozorované rozdíly je použít k testování zajímavých hypotéz – např. té, podle níž ostrovy a podobné izolované lokality usnadňují rapidní radiace, neboť znesnadňují genový tok a poskytují kolonizátorům nový ekologický prostor k zaplnění. Data to skutečně potvrzují: vyloučíme-li ptáky žijící na otevřeném moři, zjistíme, že taxony, jejichž oblast výskytu se více než z půlky rozkládá na ostrovech, mají průměrné DR daleko vyšší než ptáci převážně pevninští (0,203 oproti 0,153). Řada ostrovů vykazuje neobvykle vysoké rychlosti diverzifikace – především to platí pro pěvce v Karibiku, severním Atlantiku a Pacifiku (např. opět kruhoočkovití).

Geografická variace v druhově-specifických rychlostech diverzifikace a oblasti s nejvyšším zastoupením druhů s vysokým DR. Mapy jsou rozděleny na čtverce o velikosti 110 × 110 km, přičemž obarvení každého políčka je určeno průměrem DR hodnot pro všechny ptačí druhy vyskytující se v daném čtverci. Průměr je vážený převrácenou hodnotou velikosti areálu, což omezuje zkreslující vliv druhů s širokou oblastí výskytu. (a) všichni moderní ptáci, (b) nepěvčí ptáci, (c) pěvci. Mapy v pravém sloupci udávají, (d) jak se na diverzitě ptáků v tom kterém čtverci podílí 25% druhů s nejvyššími rychlostmi diverzifikace (DR 0,243 druhů/milion let), nebo absolutní počet takových druhů (e). Pro srovnání je zobrazen i absolutní počet všech druhů (f). (Zdroj: Jetz et al. 2012: Figure 3)
Fylogeneticky vzato se ukazuje, že pěvci mají vyšší průměrnou rychlost diverzifikace než ostatní ptáci (0,179 oproti 0,131), což by, jak autoři podotýkají, nemělo překvapovat už jen proto, že jejich prostě víc (z celkových 9993 druhů je 5966 pěvců). Toto zjištění odpovídá i skutečnosti, že jeden z posunů v tempu diverzifikace MEDUSA lokalizovala právě na bázi pěvců, konkrétně uzel Eupasseres (sensu Mayr 2005, 2011: zpěvní [Oscines] + křikaví [Suboscines] neboli všichni pěvci mimo pokřovníkovitých [Acanthisittidae]). Vysoké rychlosti diverzifikace pozorované v některých částech světa tedy táhnou především pěvci, ačkoli je pravda, že pěvci i ostatní ptáci pokrývají široké spektrum diverzifikačních rychlostí a obě skupiny také vykazují zvýšené tempo na ostrovech. Jetz et al. (2012) přesto odhalují zajímavý efekt: vyšší rychlosti diverzifikace v Asii a Severní Americe táhnou primárně nepěvčí ptáci (racci, datli, Aequornithes), zatímco na vysokém průměrném tempu diverzifikace v Jižní Americe a především pak jižní části hlavního pásma deštných lesů se podílejí prakticky výhradně radiace pěvců, hlavně hrnčiříků a klouzálků (vytvářejících klad označovaný buď jako Furnariidae sensu lato, nebo štěpený do "čeledí" Dendrocolaptidae, Scleruridae a Furnariidae sensu Moyle et al. 2009b). Autoři tento vzorec vysvětlují zvýšením plochy oblastí s mírným podnebím v posledních 10 až 20 milionech let a v nedávné době i ústupem zalednění ve vyšších zeměpisných šířkách.
Tato zjištění kontrastují s nedávnými studiemi, podle níž rychlosti speciace i vymírání výrazně vzrůstají se zeměpisnou šířkou (Weir & Schluter 2007). Přestože většina druhů v horním kvartilu rychlosti diverzifikace má skutečně střední zeměpisnou šířku svého areálu mezi 30° severní a 30° jižní šířky, totéž platí pro všechny druhy (tropy jsou prostě co do počtu taxonů bohatší) a v nížinatých tropech je co doo počtu překonávají pomalu diverzifikující druhy. Průměrná rychlosti diverzifikace nevykazuje ve vztahu k zeměpisné šířce žádný výrazný trend a není signifikantně odlišná ani pro intra- verzus extra-tropické střední zeměpisné šířky. Rozdíly v diverzitě na různých zeměpisných šířkách tak nelze vysvětlit prostými rozdíly v celkové rychlosti diverzifikace (tedy že by v tropech měla speciace "náskok" před extinkcí), contra Weir & Schluter (2007). Potvrzena se tím naopak zdá být hypotéza, že větší diverzita v tropech je způsobena delší dobou existence a větší rozlohou deštných lesů oproti jiným prostředím (Jetz & Fine 2012) – jak připomíná Ricklefs (2012) v krátkém shrnutí a diskuzi nové práce, ještě 30 milionů let zpátky byla Země převážně tropická, než se začaly rozšiřovat oblasti mírného a boreálního klimatu. Zatímco na rovníku samotném je v průměrné rychlosti diverzifikace patrný propad, mezi 10° a 15° zeměpisné šířky hodnoty narůstají (což Jetz a spol. opět dávají do souvislosti s vyšší klimatickou dynamikou v posledních 10 milionech let) a ve vyšších šířkách opět klesají. Autoři ale upozorňují, že tato šířková variace je méně významná než variace mezi polokoulemi, mezi pevninskými a ostrovními druhy a konečně i s výskytem, rozšířením a evoluční dynamikou konkrétních ptačích kladů v tom kterém místě. Ricklefs (2012) připomíná, že absence zjevných rozdílů v tempu diverzifikace v závislosti na zeměpisné šířce byla nedávno rovněž doložena u savců (Soria-Carrasco & Castresana 2012).

Odchylky druhově-specifické rychlosti diverzifikace v závisloti na zeměpisné šířce. Každý černý bod udává DR jednoho ptačího druhu a střední zeměpisnou šířku jeho globálního areálu. Mezi rychlostí diverzifikace a střední zeměpisnou šířkou výskytu není žádný signifikantní vztah a DR nevykazuje významné rozdíly mezi intra- a extra-tropickou polohou střední zeměpisné šířky (kde intra-tropický výskyt = zeměpisná šířka 23° a menší). Černá linka udává průměrnou rychlost diverzifikace v dané zeměpisné šířce. Čárkovaná linka udává práh, nad nímž leží horní kvartil rychlostí diverzifikace (DR 0,243 druhů/milion let a vyšší). Odstín okolo teček udává oblasti grafu s vyšší hustotou druhových bodů. (Zdroj: Jetz et al. 2012: Figure 4)
6. Robustnost a nejistota
Důležité je i to, že všechny tyto výsledky jsou robustní vůči volbě topologické kostry (Ericson et al. 2006: ESM-Figure 6 vs. Hackett et al. 2008) a vůči zahrnutí druhů s nedostatkem dat. Jetz et al. (2012) v tomto místě také konstatují, že jejich strom, snažící se najít vyvážený kompromis mezi obsáhnutím zbývající nejistoty ohledně ptačí fylogeneze a integrací nejnovějších taxonomických dat, nebude použitelný pro všechny účely. Absolutní rychlosti diverzifikace se zase odvíjejí od stáří korunního kladu ptáků (Neornithes) a to zas přímo závisí na použití konkrétních kalibračních fosilií. (Odhad otištěný ve studii jej mimochodem stanovuje na ~108 Ma, což je příjemné a rozhodně realističtější než 127–131 Ma, které nedávno nahlásili Haddrath & Baker [2012].) Především v tropech se pak jistě skrývá ještě mnoho kryptických druhů, které mohou dále změnit průměrné rychlosti diverzifikace oblasti. Nakonec Jetz et al. (2012) přiznávají i omezení vyplývající z toho, že jejich studie zahrnuje jen žijící taxony a nemůže tak obsáhnout informace o kompletně vymřelých kladech typu novozélandských moa.
7. Ptačí accelerando: anomálie, nebo zákonitost skrytá pod neúplným samplingem?
Geografické rozdíly, které z dat vyvstaly teprve s první globální analýzou rozdílů v druhově-specifických rychlostech diverzifikace v prostoru i čase, se zdají být velice výrazné a naznačují, že ptáci jsou v nějakém ohledu výjimeční: jejich rostoucí tempo diverzifikace v posledních 50 milionech let je velmi zvláštní. Jetz et al. (2012) spekulují, že adaptivní "nad-zóna", do níž ptáci nejpozději od raného eocénu diverzifikují, ještě pořád nemusí být nasycena a příležitostí pro další expanzi v ní přibývá. Konzervativnější vysvětlení je, že nasycení obsazeného ekologického prostoru, indikované analýzami jiných skupin, je jen artefaktem nedostatečného samplingu. Saturace je ale pravděpodobně stále reálným fenoménem u menších kladů s omezeným geografickým a/nebo ekologickým rozsahem (Phillimore & Price 2008; Rabosky & Lovette 2008a). Jetz a spol. navrhují tyto problémy řešit porovnáváním kompletních globálních stromů, jako je ten jejich, s lokálními analýzami menších skupin. Předpovídají, že ačkoli na menších geografických a ekologických meřítkách rychlost diverzifikace skutečně s časem klesat v souladu s hustotně-závislým modelem, tyto menší vzorce se nesloží a nevyprodukují stejný vzorec na větším, globálním měřítku. Co platí jednotlivě pro budníčky, lesňáčkovité a trogony neplatí pro skupinu z nich složenou; jednotlivé adaptivní radiace mohou být ve svém rozsahu omezeny, ale na diverzitu skupiny, která je produkuje, se žádný takový limit nevztahuje. Ptačí biodiverzita je víc než součet jejích částí.
8. Pagel (2012): první kritika
Současně se studií byl v Nature otištěno ještě citované shrnutí z pera Roberta Ricklefse, který má sám na kontě působivé množství studií o vzorcích ptačí (a především pak pěvčí) biodvierzity: ukázal, které pěvčí klady jsou rozmanitější nebo méně rozmanité, než bychom očekávali, a ukázal, že anomálie mají původ spíš v biogeografii než klíčových inovacích (Ricklefs 2003); doložil korelaci mezi morfologickou variabilitou a počtem druhů, naznačující vliv kladogeneze na evoluci znaků (Ricklefs 2004), a ukázal, že druhově chudé klady lze většinou nalézt na periferii pěvčího morfoprostotu (Ricklefs 2005). Za druhým textem reagujícím na megafylogenezi Jetze a spol. stojí fylogenetik Mark Pagel. Ten zdůrazňuje, že studie typu té od Jetze a spol. jsou nesmírně cenné v tom, že umožňují rekonstruovat řadu dalších aspektů evoluce než je "holá" fylogeneze, včetně ancestrálních znakových stavů, tempa speciace nebo rychlosti morfologických změn. Vedou tedy k úplnějšímu pochopení evolučních událostí a procesů tvarujících historii skupiny, a obrovská statistická síla vyplývající ze samplingu, který je hustý i široký současně, umožňuje zamítat hypotézy s větším stupněm jistoty.
Poté ale Pagel (2012) přichází ke kritice. Upozorňuje, že spoléhání se na taxonomii při umísťování druhů nezastoupených genetickými daty a na předchozí fylogenetické studie při konstrukci topologické kostry neumožnilo dostatečně prozkoumat "the possible universe of avian relatedness" (Pagel 2012:2). Místo toho by druhy, od nichž máme molekulární data, měly mít možnost shlukovat se tak, jak diktují data (bez předem vynucených uzlů), a druhy, od nichž sekvence nemáme, by měla do stromu zařadit fylogenetická analýza jiných typů dat (morfologie, chování). Osobně si myslím, že ani jedna z těchto výtek není zvlášť užitečná; jistě je to pravda, ale nasbírat dostatečné množství non-molekulárních dat pro fylogenetickou analýzu 3330 ptáků se nezdá být příliš reálné. Stejně tak výhrada, že Jetz et al. (2012) do své studie nezahrnuli fylogenetická data z nedávné studie o zrychlené speciaci u barevně polymorfických ptáků (Hugall & Stuart-Fox 2012), která přišla s konvenční fylogenetickou analýzou zhruba 4000 druhů pěvců, je spíš jen formální. Vzhledem k tomu, že práce Hugalla a Stuart-Fox vyšla online 9. května 2012 a studie Jetze a spol. 31. října 2012, je těžké vidět, jak by ta druhá mohla zahrnout jakákoli data z té první.
Jedinou skutečně zajímavou kritiku představuje ta nejvěcnější. Většina ze závěrů Jetze a spol. je založena na jejich vlastním měřítku druhově specifického diverzifikačního tempa (DR), které je pouze převrácenou hodnotou měřírka ES, sčítajícího zlomky délek větví vedoucích od daného druhu ke kořenu stromu. Důležité je tedy spolehlivě odhadnout délky větví, a právě tam je u metod využívajících relaxovaných molekulárních hodin (jako je BEAST) potřeba velmi velká opatrnost (Venditti et al. 2010). Pagel navíc připomíná, že v tomto případě je ohromný rozsah stromu Jetze a spol. spíš dalším důvodem k obavám než výhodou: i malá chyba v odhadu délek větví může mít na tak velké fylogenezi velký dopad na konečné DR, a trendy odvozené z těchto hodnot pak nemusejí být vůbec reálné. Doporučuje, aby se souvislosti mezi klimatem, ekologií a diverzifikačními rychlostmi, které Jetz et al. (2012) předkládají, testovaly standardní komparativní metodou, tzn. hledáním dvojic nezávislých znaků, které se vyvíjejí pospolu, a na samotnou studii pohlíží spíš jako na první krůček na cestě k úplné fylogenezi ptáků a porozumění ptačí biodiverzitě.
9. V médiích
First ever family tree for all living birds reveals evolution and diversification (Science Daily) – podotýká, že globální fylogeneze se může hodit i pro stanovování priorit v konzervační biologii.
Exhaustive family tree for birds shows recent, rapid diversification (Science Daily)
Are birds accelerating their rate of evolution? (Science 2.0)
10. Acknowledgments
Děkuji Rafaelu Maiovi za zaslání kompletního PDF nové studie.
Zdroje:
- Alfaro ME, Santini F, Bock C, Alamillo H, Dornburg A, Rabosky DL, Carnevale G, Harmon LJ 2008 Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. Proc Natl Acad Sci USA 106(32): 13410–4
- Barker FK, Cibois A, Schikler P, Feinstein J, Cracraft J 2004 Phylogeny and diversification of the largest avian radiation. Proc Natl Acad Sci USA 101(30): 11040–5
- Bertelli S, Porzecanski AL 2004 Tinamou (Tinamidae) systematics: a preliminary combined analysis of morphology and molecules. Ornitol Neotrop 15(Suppl): 293–9
- Boyd JH III 2012 Taxonomy in Flux Checklist 2.66a: Core Passerida. Accessed November 3, 2012 at http://jboyd.net/Taxo/List21.html#sylvioidea
- Brown JW, van Tuinen M 2011 Evolving perceptions on the antiquity of the modern avian tree. 306–24 in Dyke GJ, Kaiser G, eds. Living Dinosaurs: The Evolutionary History of Modern Birds. London: Wiley & Sons, Ltd.
- Clements JF, Schulenberg TS, Iliff MJ, Sullivan BL, Wood CL, Roberson D 2012 The eBird/Clements checklist of birds of the world: Version 6.7. Accessed November 3, 2012 at http://www.birds.cornell.edu/clementschecklist/corrections/Oct2012
- Ericson PGP 2012 Evolution of terrestrial birds in three continents: biogeography and parallel radiations. J Biogeogr 39(5): 813–24
- Ericson PGP, Anderson CL, Britton T, Elżanowski A, Johansson US, Källersjö M, Ohlson JI, Parsons TJ, Zuccon D, Mayr G 2006 Diversification of Neoaves: Integration of molecular sequence data and fossils. Biol Lett 2(4): 543–7
- Goloboff PA, Catalano SA, Mirande JM, Szumik CA, Arias JS, Källersjö M, Farris JS 2009 Phylogenetic analysis of 73 060 taxa corroborates major eukaryotic groups. Cladistics 25(3): 211–30
- Grant PR, Grant BR 2008 How and Why Species Multiply: The Radiation of Darwin’s Finches. Princeton, NJ: Princeton Univ Press
- Hackett SJ, Kimball RT, Reddy S, Bowie RC, Braun EL, Braun MJ, Chojnowski JL, Cox WA, Han K, Harshman J, Huddleston CJ, Marks BD, Miglia KJ, Moore WS, Sheldon FH, Steadman DW, Witt CC, Yuri T 2008 A phylogenomic study of birds reveals their evolutionary history. Science 320(5884): 1763–8
- Haddrath O, Baker AJ 2012 Multiple nuclear genes and retroposons support vicariance and dispersal of the palaeognaths, and an Early Cretaceous origin of modern birds. Proc R Soc B 279(1747): 4617–25
- Harshman J 2007 Classification and phylogeny of birds. 1–35 in Jamieson BGM, ed. Reproductive Biology and Phylogeny of Birds. Enfield (NH): Science Publishers Inc.
- Hugall AF, Stuart-Fox D 2012 Accelerated speciation in colour-polymorphic birds. Nature 485(7400): 631–4
- Jetz W, Fine PVA 2012 Global gradients in vertebrate diversity predicted by historical area-productivity dynamics and contemporary environment. PLoS Biol 10: e1001292
- Jetz W, Thomas GH, Joy JB, Hartmann K, Mooers AØ 2012 The global diversity of birds in space and time. Nature doi:10.1038/nature11631
- Jønsson KA, Fabre P-H, Fritz SA, Etienne RS, Ricklefs RE, Jørgensen TB, Fjeldså J, Rahbek C, Ericson PGP, Woog F, Pasquet E, Irestedt M 2012 Ecological and evolutionary determinants for the adaptive radiation of the Madagascan vangas. Proc Natl Acad Sci USA doi:10.1073/pnas.1115835109
- Jønsson KA, Irestedt M, Fuchs J, Ericson PGP, Christidis L, Bowie RCK, Norman JA, Pasquet E, Fjeldså J 2008 Explosive avian radiations and multi-directional dispersal across Wallacea: Evidence from the Campephagidae and other Crown Corvida (Aves). Mol Phylogenet Evol 47(1): 221–36
- Mayr G 2005 The Paleogene fossil record of birds in Europe. Biol Rev 80(4): 515–42
- Mayr G 2011 Metaves, Mirandornithes, Strisores, and other novelties – a critical review of the higher-level phylogeny of neornithine birds. J Zool Syst Evol Res 49(1): 58–76
- McCormack JE, Harvey MG, Faircloth BC, Crawford NG, Glenn TC, Brumfield RT 2012 A phylogeny of birds based on over 1,500 loci collected by target enrichment and high-throughput sequencing. arXiv:1210.1604
- Morgan-Richards M, Trewick SA, Bartosch-Härlid A, Kardailsky O, Phillips MJ, McLenachan PA, Penny D 2008 Bird evolution: testing the Metaves clade with six new mitochondrial genomes. BMC Evol Biol 8: 20
- Moyle RG, Chesser RT, Brumfield RT, Tello JG, Marchese DJ, Cracraft J 2009b Phylogeny and phylogenetic classification of the antbirds, ovenbirds, woodcreepers, and allies (Aves: Passeriformes: infraorder Furnariides). Cladistics 25: 386–405
- Moyle RG, Filardi CE, Smith CE, Diamond J 2009a Explosive Pleistocene diversification and hemispheric expansion of a ‘‘great speciator’’. Proc Natl Acad Sci USA 106(6): 1863–8
- Nee S, Mooers AØ, Harvey PH 1992 Tempo and mode of evolution revealed from molecular phylogenies. Proc Natl Acad Sci USA 89(17): 8322–6
- O'Connor JK, Chiappe LM, Bell A 2011 Premodern birds: avian divergences in the Mesozoic. 39–114 in Dyke GJ, Kaiser G, eds. Living Dinosaurs: The Evolutionary History of Modern Birds. London: Wiley & Sons, Ltd.
- Pagel M 2012 First steps for birds. Nature doi:10.1038/nature11642
- Phillimore AB, Price TD 2008 Density-dependent cladogenesis in birds. PLoS Biol 6(3): e71
- Rabosky DL 2006 LASER: A maximum likelihood toolkit for detecting temporal shifts in diversification rates from molecular phylogenies. Evol Bioinform 2: 247–50
- Rabosky DL, Lovette IJ 2008a Density dependent diversification in North American wood warblers. Proc R Soc B 275(1649): 2363–71
- Rabosky DL, Lovette IJ 2008b Explosive evolutionary radiations: decreasing speciation or increasing extinction through time? Evolution 62(8): 1866–75
- Redding DW, Mooers AØ 2006 Incorporating evolutionary measures into conservation prioritization. Conserv Biol 20(6): 1670–8
- Reddy S, Driskell A, Rabosky DL, Hackett SJ, Schulenberg TS 2012 Diversification and the adaptive radiation of the vangas of Madagascar. Proc R Soc B 279(1735): 2062–71
- Ricklefs RE 2003 Global diversification rates of passerine birds. Proc R Soc B 270(1530): 2285–91
- Ricklefs RE 2004 Cladogenesis and morphological diversification in passerine birds. Nature 430(6997): 338–41
- Ricklefs RE 2005 Small clades at the periphery of passerine morphological space. Am Nat 165(6): 651–9
- Ricklefs RE 2006 Global variation in the diversification rate of passerine birds. Ecology 87(10): 2468–78
- Ricklefs RE 2012 Disconnects in diversity. Nature doi:10.1038/nature11642
- Sanderson MJ, Mahon MC, Steel M 2010 Phylogenomics with incomplete taxon coverage: the limits to inference. BMC Evol Biol 10: 155
- Sibley CG, Ahlquist JE 1990 Phylogeny and Classification of Birds: A Study in Molecular Evolution. New Haven: Yale Univ Press. 1008 p
- Soria-Carrasco V, Castresana J 2012 Diversification rates and the latitudinal gradient of diversity in mammals. Proc R Soc B 279(1745): 4148–55
- Stadler T 2011 Inferring speciation and extinction processes from extant species data. Proc Natl Acad Sci USA 108(39): 16145–6
- Suh A, Paus M, Kiefmann M, Churakov G, Franke FA, Brosius J, Kriegs JO, Schmitz J 2011 Mesozoic retroposons reveal parrots as the closest living relatives of passerine birds. Nature Comms 2: 443
- Venditti C, Meade A, Pagel M 2010 Phylogenies reveal new interpretation of speciation and the Red Queen. Nature 463(7279): 349–52
- Weir JT, Schluter D 2007 The latitudinal gradient in recent speciation and extinction rates of birds and mammals. Science 315(5818): 1574–6