Bakker a Galton ani netušili, jak šokujícím způsobem se jejich krátká větička, spíše mimochodem vložená do článku podporujícího dinosauří monofylii, v budoucnu ukáže být pravdivá. Když ji v roce '74 psali, dinosauří fylogenetika byla ještě v plenkách. Teropodi byli běžně dělení do dvou kategorií: velkotělých, masivně stavěných karnosaurů, jako byli Allosaurus a Tyrannosaurus, a drobných, spíše gracilních celurosaurů, zastoupených např. ornitolestem. Předky ptáků jsme podle Ostromových teorií, které tehdy stále byly horkou novinkou, měli hledat výhradně v druhé z těchto skupin.
Odhad obou autorů se potvrdil o třináct let později. Kurzanov (1987) ve své monografii o avimimovi nahlásil, že loketní kost tohoto zvláštního dinosaura vykazuje ulnární papily – zvláštní hrbolky, na něž se u některých žijících ptáků upínají brky sekundárních letek. Objev naneštěstí zapadl; mohu-li si zaspekulovat o tom, proč se tak stalo, bylo to nejspíš proto, že podle celé řady autorů byl Avimimus blíže příbuzný žijícím ptákům než Archaeopteryx, takže jestli měl peří, nic moc se neděje. S přímou evidencí pro přítomnost peří – totiž s otisky – navíc přišel až Sinosauropteryx (Ji & Ji 1996), který už by se za ptáka dal zaměnit jen velmi těžko. Do roku 2005 byla nějaká forma protopeří nahlášena od zástupců každé větší dinosauří skupiny, stojící na kladogramu mezi sinosauropterygem a moderními ptáky (Ji et al. 1998; Schweitzer et al. 1999; Xu et al. 1999a, b; Ji et al. 2005). Jedinou výjimkou jsou ornitomimosauři, kde se původně nahlášený vláknitý tělesný pokryv (Pérez-Moreno et al. 1994) ukázal být chybně interpretovanými svalovými vlákny (Briggs et al. 1997).
V roce 2004 přišel objev, který sice nebyl tak úplně šokující, přesto jej ale šlo pokládat za malou revoluci: Dilong paradoxus, opeřený tyrannosauroid (Xu et al. 2004). Konec konců, tyrannosauroidi jsou podle současných fylogenetických hypotéz jen o jeden uzel na kladogramu pod kladem (Sinosauropteryx + Neornithes), a posunout výskyt znaku o jeden uzel dolů zas tak velkou změnou není. Navíc Dilong nebyl žádný Tyrannosaurus: nejenže svého populárního severoamerického příbuzného předcházel o 60 milionů let (časová mezera mezi ním a tyrannosaurem byla tím pádem skoro stejně velká, jako mezi tyrannosaurem a současností), ale především ho vůbec nepřipomínal. Šlo o gracilní zvíře dlouhé pouhých 1,6 m. Média pochopitelně ihned zaplnily spekulace o opeřeném T-rexovi, které Xu s kolegy podpořili jen napůl:
Jsme tedy zpátky tam, kde jsme byli: překážkou pro představu opeřeného tyrannosaura není fylogeneze, nýbrž velikost. Jsme zvyklí na to, že savci podobné velikosti (sloni, nosorožci – nemluvě o menších kytovcích, kteří jsou ale trochu jiný případ) už na sobě moc srsti nemají. Největší dosud známý opeřený dinosaur, terizinosaur
Beipiaosaurus, měřil 2,2 m a vážil necelých 90 kg. Osmimetrový, jedenapůltunový opeřený
Allosaurus, jakého v roce 1975, uprostřed nadšení dinosauří renezance, nakreslil
Greg Paul, se tedy zdál být značně nepravděpodobný.
Až do minulé středy.
2. "What is big, fluffy, and could tear you to shreds?"
Odpověď zní:
Yutyrannus huali, 9 m dlouhý opeřený tyrannosauroid popsaný v
Nature týmem soustředěným okolo slavného paleontologa Xu Xinga, který si na liaoningských opeřencích postavil kariéru. Geniální název podkapitolky jsem si vypůjčil
od Thomase Holtze, který s ní uvodil oznámení o objevu na Dinosaur Mailing Listu.
Fotografie lebky exempláře ELDM V1001, paratypu a nejmenšího známého jedince taxonu Yutyrannus huali. (Zdroj: archosaurmusings.wordpress.com)

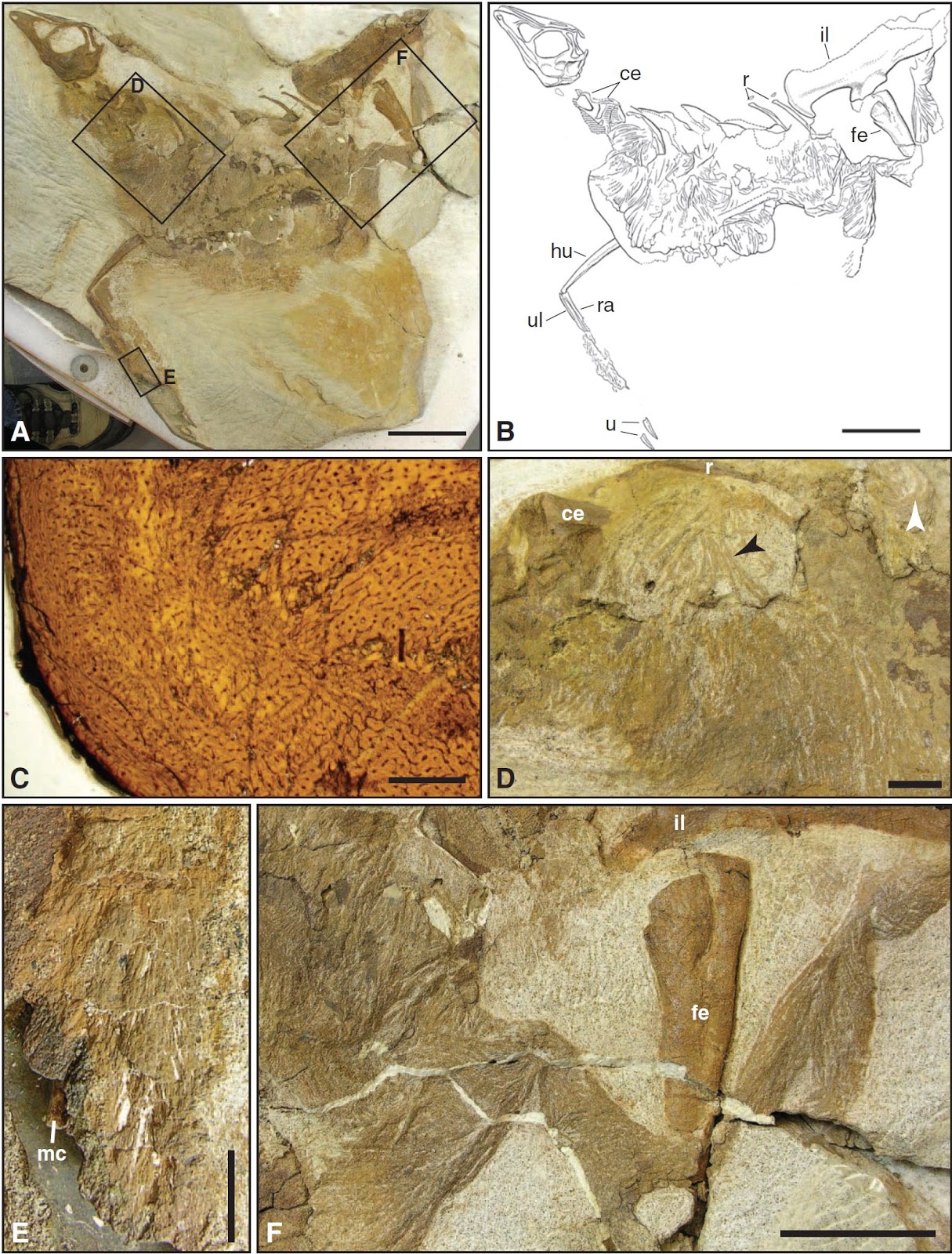
Interpretační kresby koster ZCDM V5000 (holotyp) a ZCDM V5001 (paratyp), odkrytých z jediného bloku horniny, a lebky ELDM V1001, zobrazené výše. Zkratky: (aop) přídatný očnicový výběžek, (-c) krkavčí kost, (cav) ocasní obratle, (cev) krční obratle, (clp) kultriformní výběžek = parasfenoidní rostrum, (co) výduť, (cp) rohu podobný výběžek kosti slzné, (dr) hřbetní žebra, (dv) hřbetní obratle, (-ec) zevní kost křídlová, (ga) břišní žebra, (-fe) stehenní kost, (-fi) lýtková kost, (g) rýha, (-h) pažní kost, (-il) kyčelní kost, (-is) sedací kost, (l-) levý, (-m) ruka, (ma) dolní čelist, (mf) maxillární okno, (np) výstupky na nosní kosti, (-p) chodidlo, (-pa) patrová kost, (pnr) pneumatické výklenky, (pu) stydká kost, (-r) vřetenní kost, (r-) pravý, (ri) hřeben, (-s) lopatka, (sk) lebka, (sp) podočnicový výběžek, (-sq) šupinová kost, (sr) hřeben na nadúhlové kosti, (sy) synsakrum, (-t) tibiotarzus, (-u) loketní kost. Platnost pruhu měřítka se vztahuje pouze na horní část obrázku. (Modifikováno z Xu et al. 2012: Figure 1 a Figure 2)
2.1 Obecná fakta
Nový taxon byl popsán na základě téměř kompletní kostry odkryté ze spodnokřídových vrstev souvrství Yixian v čínské provincii Liaoning. Kromě ní mají paleontologové k dispozici ještě dva paratypy: jeden exemplář je kompletní, druhý bez ocasu, oba však plně artikulované. Na základě rovnice, kterou předložili Christiansen & Fariña (2004), byla hmotnost typového exempláře spočtena na 1414 kg, zatímco majitelé dvou dalších koster vážili pouze 596 a 493 kg. Spolu s několika kosterními znaky (jako jsou např. stále otevřené švy mezi obratlovými centry a neurálními oblouky) tento velikostní rozdíl svědčí o tom, že oba paratypy patří nedospělým jedincům. Na základě dat, která o tyrannosauroidním růstu nashromáždili Erickson et al. (2004), Xu a kolektiv odhadují, že nejmenší exemplář byl v době smrti minimálně o 8 let mladší než holotyp.
Prenomen ("rodové jméno"), který Xu et al. (2012) zvolili, kombinuje latinské slovo pro "tyrana" s mandarínským výrazem pro "peří" (
yu), "druhové jméno" znamená v mandarínštině "krásný" – vzhledem ke kvalitě fosilií jde o přízvisko vskutku případné. Diagnóza se zaměřuje hlavně na odlišnosti od ostatních tyrannosauroidů a především od siamotyranna, jediného tyrannosauroida srovnatelné velikosti známého z dané oblasti. Autapomorfickým (čili jedinečným a současně odvozeným) znakem je však sagitální hřeben formovaný nosními a předčelistními kostmi, disponující zdrsnělým povrchem, mnoha otvůrky (vedoucími zřejmě do vnitřních, vzduchem vyplněných prostor) a výčnělky na horním okraji. Podobné útvary vidíme u guanlonga, zástupce bazální tyrannosauroidní skupiny
Proceratosauridae, a již zmiňovaného karcharodontosaura konkavenatora; všichni svrchnokřídoví tyrannosauroidi dále vykazují na vrchní straně lebky nějaké zdrsněliny, které by s částí jutyrannova hřebenu mohly být homologické.
Autoři poznamenávají, že mezi jednotlivými exempláři existují určité odlišnosti, z nichž většina je pravděpodobně ontogenetického původu. Srovnáváním menších a větších jedinců dospěli Xu a spol. k závěru, že s rostoucím věkem se lebka stávala proporčně kratší a vyšší, stejně jako předčelistní kost. Jak zvíře stárlo, maxillární okno se posouvalo více dopředu. Většina takto proměnlivých znaků se nepřekvapivě vyskytuje na lebce; exempláře se však liší např. i tvarem pánevních kostí. Takový rozdíl se dá ontogeneticky interpretovat podstatně hůře – snad odráží spíše pohlavní dimorfizmus. Xu et al. (2012) se také pokusili o morfometrický rozbor fosilií, který by dokázal odkrýt další změny provázející růst zvířete. Jak autoři objasňují v
doplňkových informacích volně dostupných online, jeho provedení bylo do značné míry komplikováno malým vzorkem (2 exempláře jsou navíc skoro stejně velké) a omezeným počtem elementů přítomných u všech třech koster. Přesto z něj však šlo cosi vyvodit: jak zvíře rostlo, lopatka a kyčelní kost byly v poměru ke kosti stehenní čím dál tím menší a menší, zatímco u tyrannosauridů zůstával poměr mezi velikostí kosti kyčelní a femuru po celou ontogenezi skoro stejný a lopatka se dokonce proporčně zvětšovala (Currie 2003).
Jedna z mnoha ukázek úžasné kvality, v níž se jutyrannův kosterní materiál dochoval: detail na přední končetinu (c) a pánev spolu se zadní končetinou (d) exempláře ZCDM V5001. Za povšimnutí stojí pleziomorfická přední končetina, která je oproti svrchnokřídovým tyrannosauridům poměrně dlouhá a vybavená ještě třemi funkčními prsty namísto dvou. (Zdroj: Xu et al. 2012: Figure S2)
Yutyrannus vykazuje několik primitivních (pleziomorfických) znaků, které naznačují, že ač patří mezi tyrannosauroidy, nespadá do užšího kladu
Tyrannosauridae, kde se podobně velcí predátoři vyskytovali hlavně. Jde např. o vzadu posazené oválné otvory pro nozdry nebo rýhu na horní čelisti rovnoběžnou se spodním okrajem předočnicové jamky. Xu et al. (2012) zmiňují, že některé z těchto pleziomorfií byly dřívějšími fylogenetickými analýzami odhaleny coby sjednocující znaky bazálně-tyrannosauroidního kladu
Proceratosauridae (Rauhut et al. 2009), jehož odhalení jsem zmínil jako jeden z
paleontologických úspěchů roku 2009. Jejich analýzy však tuto interpretaci nepodporují a údajné proceratosauridní synapomorfie optimizují jako společné znaky všech tyrannosauroidů.
Na druhou stranu nový taxon vykazuje rysy, které ho k asijským a severoamerickým vrcholovým predátorům typu tyrannosaura, tarbosaura, albertosaura nebo gorgosaura řadí blíž než dilonga, guanlonga nebo proceratosaura. Sem patří velká, vysoká lebka; vysoká a masivní předčelistní kost; velký, rohům podobný kuželovitý výběžek vyrůstající z horního konce slzných kostí; nebo malé vnější mandibulární okno. Směsku primitivních a odvozených znaků vykazuje také postkraniální kostra. Pneumatizace otvorů např. nedosahuje míry viděné u odvozených tyrannosauridů, ale na neurálních trnech krčních obratlů už vidíme velké obruby pro připojení meziobratlových vazů, které se zřejmě podílely na podpírání masivní hlavy. Zadní končetiny se opět podobaly spíše bazálním tyrannosauroidům (nebo karnosaurům, jako byli
Allosaurus či
Carcharodontosaurus) než odvozeným tyrannosauridům.
Vzhledem k této kombinaci znakových stavů nijak nepřekvapí, že na kladogramu
Yutyrannus skutečně stojí "v půli cesty" mezi bazálními tyrannosauroidy a tyrannosauridy. Na tomto výsledku se shodly dvě nezávislé fylogenetické analýzy (každá postavená na jiné matrici), které Xu et al. (2012) k rozřešení fylogenetické pozice taxonu použili. Nejprve byl
Yutyrannus dosazen do datasetu z popisu haplocheira (Choiniere et al. 2010), který představuje jednu z novějších iterací TWG (Theropod Working Group) matrice. Ten disponuje širokým vzorkem neptačích teropodů, tyrannosauroidů ale obsahuje poměrně málo (bez jutyranna 6). Dobrý byl tak hlavně k tomu, aby ukázal, že nový čínský taxon skutečně patří k tyrannosauroidům a ne jiným dravým teropodům, což vzhledem k některým zvláštním znakům nemusí být zcela evidentní (viz níže).
Pozici v rámci tyrannosauroidů bylo potom možné určit až s pomocí jemnějšího rozboru. Xu a jeho tým pro tento účel vybrali datovou matrici z Brusatte et al. (2010), jejíž potenciál dále využili hned dvakrát: v jedné dílčí analýze do něj dosadili jutyranna coby jedinou operační taxonomickou jednotku (OTU – to, co na výsledném stromu stojí na špičkách větví), v druhé udělili samostatné OTU každému z tří exemplářů. Tím zároveň otestovali hypotézu, že všechny tři kostry skutečně patří jednomu a témuž taxonu: test dopadl pro hypotézu příznivě; všechny tři kostry skutečně tvoří monofyl vylučující všechny ostatní tyrannosauroidy. Kódování všech jutyrannů odděleně navíc rozřešilo výsledný strom lépe než analýza s jediným OTU, díky čemuž se na výsledný strom vrátili např. monofyletičtí proceratosauridi složení z proceratosaura, guanlonga a sinotyranna (viz níže).

Fylogeneze tyrannosauroidů; Yutyrannus je ten ochmýřený fešák vpravo uprostřed. Vlevo: striktní konsenzus 3 nejúspornějších stromů o celkové délce 629 kroků, vzniklých dosazením tří exemplářů jutyranna každého zvlášť do datové matrice Brusatteho a spol. (2010). Vpravo osekaná verze téhož stromu kalibrovaná podle časové osy. Byl jsem původně v pokušení napsat, že Yutyrannus vyvinul své gigantické rozměry konvergentně na svrchnokřídových tyrannosauridech a jejich nejbližších příbuzných, podrobnější pohled na strom ale odhalí, že velikostní evoluce tyrannosauroidů byla víc než cokoli jiného matoucí. Pomineme-li podivné konvergentní nabytí velmi velkých tělesných rozměrů u proceratosauridů v podobě 10 m dlouhého sinotyranna, stále se nám malé a velké taxony podivným způsobem střídají. Třímetrový Raptorex je sesterskou skupinou kladu (Appalachiosaurus + Tyrannosauridae), složeného ze zvířat s délkou 7 m a větší. Aby toho nebylo málo, ke kladu (Raptorex + Tyrannosauridae) stojí v sesterském vztahu opět asi sedmimetrový Dryptosaurus. Takový rozpor lze vysvětlit, přijmeme-li tvrzení, že Raptorex je jen mládětem tarbosaura (Fowler et al. 2011) a nikoli ukázkou skromných začátků tyrannosauridího tělního plánu (Sereno et al. 2009). I pak nám ale zbývá k vysvětlení fakt, že do mezery mezi dvěma velkotělými taxony – jutyrannem a kladem (Dryptosaurus + Tyrannosauridae) – vklouzly dva taxony nepřesahující 4 metry. Otázka, kolikrát tyrannosauroidi gigantizmus vyvinuli a zase druhotně ztratili, se tedy nezdá být zrovna triviální. (Modifikováno z Xu et al. 2012: Figure 3 a Figure S6)
2.2 Peří
To nejúžasnější na novém čínském objevu však nejsou tři téměř kompletní, artikulované kostry gigantického spodnokřídového tyrannosauroida v různých ontogenetických stadiích. Jsou jím tři téměř kompletní, artikulované kostry gigantického spodnokřídového tyrannosauroida
s protopeřím.
Tenká vlákna lze skutečně zaznamenat kolem všech 3 fosilií. U holotypu jsou patrná v zadní části ocasu, jsou vzájemně rovnoběžná a s podélnou osou ocasu svírají úhel asi 30°. U ZDCM V5001 obklopují pánevní oblast a metatarzály, u ELDM V1001 horní stranu krku a dlouhou kost, kterou autoři odhadují na humerus. Zdá se tedy, že opeření pokrývalo větší část těla. Ve všech případech jsou otisky nahromaděné u sebe tak hustě, že nelze rozeznat detaily jejich morfologie. Mohlo by jít o drobná vlákna podobná těm dilongovým nebo sinosauropterygovým (která jsou ohebná a jen asi 0,1 až 0,2 mm tenká; Xu et al. 2009), ale také o EBFFs, "Elongated Broad Filamentous Feathers", jaká byla tři roky zpátky nahlášena od beipiaosaura (Xu et al. 2009). Tento zvláštní typ peří je spíše tuhý, štětinovitý, daleko širší (2 až 3 mm) a vykazuje mimořádnou podobnost jak se štětinami ptakopánvých, tak i "srstí" pterosaurů. Mohlo by dokonce jít i o pokročilejší formy peří s vícero vlákny vyrůstajícími z jedné společné báze – kvalita fosilií to prostě neumožňuje zjistit. Délka filamentů začíná na 15 a končí minimálně na 20 centimetrech. Jsou-li skutečně nevětvené – což se zdá jako rozumný předpoklad, jelikož nic jiného než samostatná nevětvená vlákna z tohoto místa kladogramu s jistotou neznáme (Xu & Guo 2009) – nápadně se podobají pernatému pokryvu dnešních emu a kasuárů, což ospravedlňuje můj titulek. (Hurá! Žádná klamavá reklama!) S tímto postřehem mě ovšem
na DML předběhl Thomas Holtz.
(Tato podobnost je vlastně tak nápadná, že má smysl se ptát, zda nazývání jutyrannova integumentu jiným jménem, totiž jako "protopeří", zbytečně nezastírá shodnou podstatu obou struktur. V této věci nemám žádné vyhraněné preference. Na jednu stranu by šlo argumentovat, že peří kasuárů a emu je výsledkem druhotného zjednodušení, zatímco u tyrannosauroidů byla tato jeho podoba prvotní, pleziomorfická. To na druhou stranu neznamená, že by oba tělní pokryvy nemohly vznikat v důsledku stejných developmentálních procesů. Co hůř, onen příjemně primitivní, vláknitý zjev "protopeří" může být pouhým tafonomickým artefaktem. Čížek lesní rozmáčknutý v tiskařském lisu
je také ve výsledku obklopen halem krásně vláknitého integumentu, přestože v normálním stavu disponuje úplně normálním peřím s ostnem, větvemi a háčky [Foth 2011]. Proces fosilizace se rozmáčknutí lisem v lecčems podobá.)
Detail na zadní část ocasu u jutyrannova holotypu ZCDM V5000, zobrazující nádherně zachovalé protopeří. (Zdroj: archosaurmusings.wordpress.com)
Peří u ELDM V1001. Fotografie zachycují otisky vláknitého integumentu nad krkem (a), detail na téže místo (b) a podél polámané kosti, která se na základě velikosti zdá být kostí pažní (c). (Modifikováno z Xu et al. 2012: Figure S3)
Zatímco peří na tyrannosauroidech už by dnes nikoho šokovat nemělo (viz výše), peří na zvířeti 9 m dlouhém a 1,4 tuny těžkém se přece jen nečekalo. Největší prokazatelně opeření neptačí dinosauři, kompsognátid
Sinocalliopteryx a o něco kratší, ale masivnější
Beipiaosaurus (Xu et al. 1999a, 2009), ve skutečnosti co do velikosti zaostávali nejen za ptačími giganty typu vyhynulých sloních ptáků či dromornitidů, ale i za žijícím pštrosem dvouprstým.
Yutyrannus ale beipiaosaura co do hmotnosti překonává víc než 40×, sloní ptáky pak asi 3×. Problém byl asi hlavně v tom, že neznáme žádného ptáka o hmotnosti vyšší než půl tuny; u všeho většího jsme tedy byli odkázáni na analogie se savci. Ti jsou většinou bezsrstí: díky příznivému poměru mezi objemem a povrchem těla žádnou tepelnou izolaci nepotřebují (více jsem o tom na blogu psal
v souvislosti s kolibříky). Naopak se jim hodí spíš přídavné plochy, přes které by mohli teplo ztrácet – viz sloní uši.
V chladnějších oblastech si ale i velmi velcí savci svou srst ponechají víc než rádi: mamuti, srstnatí nosorožci i bizoni by mohli vyprávět. Mohla mít opeřenost jutyranna stejnou příčinu? Podle Xu'a a kolektivu možná ano. V době, kdy území dnešní západní části provincie Liaoning obýval
Yutyrannus, se zde průměrná roční teplota vzduchu pohybovala okolo 10°C (Amiot et al. 2011), což je o 8°C méně, než bylo ve stejných zeměpisných šířkách běžné ve svrchní křídě. Většina svrchnokřídových gigantů zřejmě žila v teplém klimatu a žádnou tepelnou izolaci tedy nepotřebovala, ačkoli Xu et al. (2012) uznávají, že populace tyrannosauridů obývaly i Aljašku. Od gigantických svrchnokřídových tyrannosauridů známe otisky šupin, ne však otisky peří. To sice nemusí mnoho znamenat, jelikož peří a šupiny mohou na těle jednoho zvířete v klidu a míru koexistovat (jak ukazují nohy prakticky kteréhokoli žijícího ptáka), pokud by ale
Tyrannosaurus rex přece jen peří měl, museli by se tyrannosauroidi od savců co do vztahu mezi integumentem a velikostí dost výrazně lišit (viz však níže). Podle autorů je možné i to, že rozsah pernatého pokryvu těla byl v tyrannosauroidní evoluci velice proměnlivý a ovlivňovaly ho jak tělesné rozměry, tak i teplota okolního prostředí. V poslední řadě nelze zcela vyloučit možnost, že
Yutyrannus měl místo souvislého ochmýření po těle rozeseto jen několik drobných opeřených flíčků. Pak by byla pravděpodobnější nějaká jiná, s termoregulací nesouvisející funkce. Vhodným kandidátem by byla třeba ornamentace, která je zřejmě ze všech funkcí ptačího peří nejpůvodnější (Xu & Guo 2009).
Pro opeřeného T-rexe ovšem argumentuje Thomas Holtz Jr., který své myšlenky poprvé vyjádřil na DML a rozvedl v
komentáři pro National Geographic News a na různých blozích (
Archosaur Musings,
Tetrapod Zoology). Někteří gigantičtí savci si totiž zachovali důkladné osrstění i přesto, že žili v tropech: příkladem může být
Megatherium, čtyřtunový jihoamerický pozemní lenochod vyhubený lidským lovem ke konci poslední doby ledové. Ani vymřelí moa z Nového Zélandu či sloní ptáci z Madagaskaru konec konců neobývali zrovna mrazivé pustiny. Holtz dále upozorňuje, že
Tyrannosaurus se mohl vyskytovat i za polárním kruhem, a pak by bylo trochu divné předpokládat, že (zatím hypotetická) populace žijící na území dnešní Aljašky peří měla, zatímco ta z Nového Mexika už ne:
"Tigers live in the forests of Siberia down to the jungles of southeast Asia. It's true that Siberian tigers have thicker fur, but they're still furry down in the south."
— Thomas R. Holtz Jr. pro nationalgeographic.com
Zdá se tedy docela klidně možné (ne-li pravděpodobnější), že i
Tyrannosaurus a další svrchnokřídoví tyrannosauridi peří měli.
3. Přidaná hodnota: blogosféra
Úžasnou věcí na pan-avianní paleontologii je množství technických blogů, které píší, čtou a navzájem komentují profesionální paleontologové. Nevím, zda něčím podobným disponují dejme tomu astrofyzici, ale u dinosaurů se celý systém rozmohl natolik, že začíná konkurovat samotné primární literatuře. Však už v ní také některé blogové příspěvky
byly citovány – konec konců, blog je rigoróznějším zdrojem než "pers. comm." (= slyšel jsem to), "pers. obs." (= viděl jsem to) či "unpubl. data" (= vyšlo mi to tak, musíte mi to věřit), které jsou v odborných periodikách zcela běžné. Systém, kde paleontolog napíše svou myšlenku na web a jiný paleontolog mu ji okomentuje (ať už příznivě nebo nepříznivě), se nijak zásadně neliší od recenzního řízení, je však rychlejší a flexibilnější. Eric Buffetaut samrukii poprvé označil za pterosaura v
komentáři pod Naishovým blogem, recenzovaná studie na stejné téma mu vyšla o dva měsíce později. V jiných případech samotný článek představuje plnohodnotný výzkum, schopný bez problémů stát sám za sebe: sem spadá třeba reinterpretace konkavenatorových "ulnárních papil" coby mezisvalových linií nebo fylogenetické analýzy Andrey Caua, založené na jeho Megamatrici.
Krásnou ukázkou potenciálu profesionální blogosféry je i
Yutyrannus. Přestože drobné pochyby o tyrannosauroidním statusu nového taxonu vyjádřil už Holtz ve zde opakovaně citovaném komentáři na DML, vše pořádně začalo, až když o jutyrannovi napsal Darren Naish
na Tetrapod Zoology. Naish je na bazální tyrannosauroidy expert: byl spoluautorem při popisu eotyranna (Hutt et al. 2001), jeho osteologií se zabýval ve své doktorské práci (Naish 2006) a v současnosti o něm sepisuje monografii. Aby toho nebylo málo, Naish práci Xu'a a spol. recenzoval a jeho pohled tedy není radno brát na lehkou váhu.
O co jde? Naishovi totiž
Yutyrannus spíše než tyrannosauroida připomíná zástupce jiné skupiny velkotělých dravých dinosaurů, totiž karcharodontosaurů (
Carcharodontosauridae +
Neovenatoridae). Na blogu píše, že Xu s kolektivem sice vzali karcharodontosauří znaky jutyranna na vědomí a analýza jim ho stále vyhodila jako tyrannosauroida, současně ale vyjmenovává celou řadu podobností s konkavenatorem – taxonem, který do ani jedné z dílčích v původní publikaci zahrnut nebyl. D. Naish sám dodává, že tahle podobnost je trochu paradoxní: zvíře, o které se všichni zajímají kvůli jeho peří, by mělo být blízce příbuzné zvířeti, které... údajně mělo mít peří, ale nejspíš ho nemělo (viz výše). Podobnosti jsou však vskutku zarážející:
Concavenator stejně jako
Yutyrannus vykazuje hřeben na nosních kostech s kruhovými až eliptickými pneumatickými výklenky po stranách, navíc vykazuje několik znaků, které Xu et al. (2012) označili za tyrannosauroidní. Na jednu stranu by opeřený karcharodontosaur byl ještě o něco úžasnější než opeřený tyrannosauroid (toho už přece jen máme, bez ohledu na podivný výsledek Leeho a Worthyho; zato
Concavenator paleontologickou komunitu o opeřených karcharodontosaurech v žádném případě nepřesvědčil), na stranu druhou by nový objev zhořkl pachutí fylogenetické nejistoty* a vyvstala by nutnost čekat na to, až bude
Yutyrannus zahrnut "within a larger and more comprehensive data set" (D. Naish).
Detail na lebku jutyranna (vlevo) a konkavenatora (vpravo). Ačkoli by oba taxony měli patřit ke dvěma odlišným liniím dravých dinosaurů, vykazují některé nápadné podobnosti, jako je třeba podélný hřeben na nosních kostech s četnými výklenky, patrně pneumatickými; nebo výběžek postorbitálu zasahující do očnice. Jde o konvergence, nebo homologie? (Modifikováno z archosaurmusings.wordpress.com a Ortega et al. 2010: Suppementary Picture 1)
Příběh má ale šťastný konec. Který dataset je totiž větší než Megamatrice italského paleontologa Andrey Caua? S téměř 1400 morfologickými znaky kódovanými pro více než 360 taxonů (
stav k říjnu 2011, dnes je určitě ještě větší) jde o zdaleka nejrozsáhlejší podklad pro fylogenetickou analýzu teropodů, vedle nějž je i populární TWG matrice než pouhým trpaslíkem. A Cau, naštěstí pro nás, čte Tetrapod Zoology a současně sám bloguje. Doplnil tedy Megamatrici několika novými znakovými stavy, přidal znaky, v nichž si je
Yutyrannus podobný s konkavenatorem a projel výsledek PAUPem. Ukázalo se, že závěry Xu'a a spol. jsou skutečně robustní: nejenže Megamatrice odhalila konkavenatora jako karcharodontosaura a jutyranna coby tyrannosauroida, ale přesunutí gigantického opeřence ke konkavenatorovi vyžadovalo plných 34 kroků navíc oproti nejparsimonnějšímu stromu, což je statisticky signifikantní rozdíl.
Pravděpodobnost, že je takový rozdíl čistě náhodný, totiž činí pouhých 0,7%; daleko méně, než obvyklá pětiprocentní hladina významnosti. Daleko lehčí by paradoxně bylo přesunutí konkavenatora a jemu podobného becklespinaxe do tyrannosauroidů za jutyrannem, i tak by to ale vyžadovalo statisticky signifikantní rozdíl 16 kroků. ("Krok" v tomto případě znamená přechod mezi dvěma různými stavy jednoho znaku.) Pozice jutyranna v rámci tyrannosauroidů podle Cauovy analýzy také docela dobře odpovídá tomu, co odhalili Xu a spol.: oba rozbory se shodnou na tom, že čínský gigant je odvozenější (= blíž tyrannosauridům) než
Stokesosaurus,
Dilong,
Kileskus či proceratosauridi; jediný rozdíl panuje v tom, že dle Caua má ke svrchnokřídovým taxonům blíž než
Xiongguanlong a
Eotyrannus, zatímco podle původní publikace je tomu naopak (viz stromek výše). Zájemcům o detaily nelze než doporučit
původní Cauův článek (v italštině; dostupná je možnost automatického překladu).
*Pan-aviani ve mně probouzejí jen to nejhorší, v tomto případě např. sklony k poezii.
Zdroje:
- http://dml.cmnh.org/2012Apr/msg00056.html
- http://archosaurmusings.wordpress.com/.../the-giant-feathered-tyrannosaur...
- http://news.nationalgeographic.com/news/...yutyrannus-feathers-dinosaur...biggest/
- http://blogs.scientificamerican.com/tetrapod-zoology/.../giant-feathered-tyrannosaurs/
- http://theropoda.blogspot.com/2011/10/pregiudizi-grafici-e-la-struttura-di.html
- http://theropoda.blogspot.com/2012/04/yutyrannus-huali-xu-et-al-2012-un_06.html
- Amiot R, Wang X, Zhou Z-H, Wang X-L, Buffetaut E, Lécuyer C, Ding Z-L, Fluteau F, Hibino T, Kusuhashi N, Mo J-Y, Suteethorn V, Wang Y-Q, Xu X, Zhang F-S 2011 Oxygen isotopes of east Asian dinosaurs reveal exceptionally cold Early Cretaceous climates. Proc Natl Acad Sci USA 108: 5179–83
- Bakker RT, Galton P 1974 Dinosaur monophyly and a new class of vertebrates. Nature 248(5444): 168–72
- Briggs DEG, Wilby PR, Pérez-Moreno BP, Sanz JL, Fregenal-Martinez M 1997 The mineralization of dinosaur soft tissue in the Lower Cretaceous of Las Hoyas, Spain. J Geol Soc Lond 154: 587–8
- Brusatte SL, Norell MA, Carr TD, Erickson GM, Hutchinson JR, Balanoff AM, Bever GS, Choiniere JN, Makovicky PJ, Xu X 2010 Tyrannosaur paleobiology: new research on ancient exemplar organisms. Science 329(5998): 1481–5
- Choiniere JN, Xu X, Clark JM, Forster CA, Guo Y, Han F-L 2010 A basal alvarezsauroid theropod from the Early Late Jurassic of Xinjiang, China. Science 327(5965): 571–4
- Christiansen P, Fariña RA 2004 Mass prediction in theropod dinosaurs. Hist Biol 16: 85–92
- Currie PJ 2003 Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the Upper Cretaceous of North America and Asia. Can J Earth Sci 40(4): 651–65
- Erickson GM, Makovicky PJ, Currie PJ, Norell MA, Yerby SA, Brochu CA 2004 Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430(7001): 772–5
- Foth C 2011 On the identification of feather structures in stem-line representatives of birds: evidence from fossils and actuopalaeontology. Paläont Z 86(1): 91–102
- Fowler DW, Woodward HN, Freedman EA, Larson PL, Horner JR 2011 Reanalysis of “Raptorex kriegsteini”: A juvenile tyrannosaurid dinosaur from Mongolia. PLoS ONE 6(6): e21376
- Gierliński G 1996 Feather-like impressions in a theropod resting trace from the Lower Jurassic of Massachusetts. Mus N Arizona Bull 60: 179–84
- Hutt S, Naish D, Martill DM, Barker MJ, Newbery P 2001 A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England. Cretac Res 22(2): 227–42
- Ji Q, Currie PJ, Norell MA, Ji S-A 1998 Two feathered dinosaurs from northeastern China. Nature 393(6687): 753–61
- Ji Q, Ji S-A 1996 [On the discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds.] Chin Geol 10(233): 30–3 [in Chinese, transl. by Downs W]
- Ji Q, Ji S-A, Lü J-C, You H-L, Liu Y-Q, Liu Y-X 2005 First avialan bird from China (Jinfengopteryx elegans gen. et sp. nov.). Geol Bull China 24(3): 197–205
- Kundrát M 2004 When did theropods become feathered?—Evidence for pre-Archaeopteryx feathery appendages. J Exp Zool (Mol Dev Evol) 302B: 355–64
- Kurzanov SM 1987 [Avimimidae and the problem of the origin of birds.] Trudy Sovm Sov-Mong Paleontol Eksped 31: 5–92 [in Russian]
- Lee MSY, Worthy TH 2011 Likelihood reinstates Archaeopteryx as a primitive bird. Biol Lett 8(2): 299–303
- Naish D 2006 The osteology and affinities of Eotyrannus lengi and other Lower Cretaceous theropod dinosaurs from England. Univ of Portsmouth: Unpubl. PhD Thesis
- Ortega F, Escaso F, Sanz JL 2010 A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain. Nature 467(7312): 203–6
- Pérez-Moreno BP, Sanz JL, Buscalioni AD, Moratalla JJ, Ortega F, Raskin-Gutman D 1994 A unique multitoothed ornithomimosaur from the Lower Cretaceous of Spain. Nature 370(6488): 363–7
- Rauhut OWM, Milner AC, Moore-Fay S 2009 Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zool J Linn Soc 158(1): 155–95
- Schweitzer MH, Watt JA, Avci R, Knapp L, Chiappe LM, Norell MA, Marshall M 1999 Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous alvarezsaurid, Shuvuuia deserti. J Exp Zool (Mol Dev Evol) 285(2): 146–57
- Sereno PC, Tan L, Brusatte SL, Kriegstein HJ, Zhao X-J, Cloward K 2009 Tyrannosaurid skeletal design first evolved at small body size. Science 326(5951): 418–22
- Xu X, Guo Y 2009 The origin and early evolution of feathers: insights from recent paleontological and neontological data. Vert PalAs 47(4): 311–29
- Xu X, Norell MA, Kuang X-W, Wang X-L, Zhao Q, Jia C-K 2004 Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature 431(7009): 680–4
- Xu X, Tang Z-L, Wang X-L 1999a A therizinosauroid dinosaur with integumentary structures from China. Nature 399(6734): 350–4
- Xu X, Wang K-B, Zhang K, Ma Q-Y, Xing L-D, Sullivan C, Hu D-Y, Cheng S-Q, Wang S 2012 A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature 484(7392): 92–5
- Xu X, Wang X-L, Wu X-C 1999b A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. Nature 401(6750): 262–6
- Xu X, Zheng X-T, You H-L 2009 A new feather type in a nonavian theropod and the early evolution of feathers. Proc Natl Acad Sci USA 106(3): 832–4


































{kind=link}
{kind=link}