Z neobvyklého úhlu pohledu se na pterosaury loni zaměřil Tokita (2014), který ve své studii analyzuje evolučně-developmentální bázi pterosauří morfologie. V úvodu studie autor připomíná, že pterosauři byli první ze tří skupin obratlovců, u kterých se vyvinula schopnost letu. Jak udávají Upchurch et al. (2014), nejstarší známý fosilní materiál pterosaurů pochází z vrstev datovaných do období pozdního karnu nebo raného noru (stáří okolo 227 milionů let) a bývá přiřazován taxonu Eudimorphodon, který byl již zjevně schopným letcem.* První dinosauři, kteří mohli být schopní nějaké formy pohybu vzduchem (nejspíše omezené na plachtění na krátké vzdálenosti) – Anchiornis a Xiaotingia (Longrich et al. 2012; Dyke et al. 2013; Sorkin 2014) – jsou přitom staří 161 milionů let (Liu et al. 2012) a první letouni – Onychonycteris a Icaronycteris – jen 52,5 milionů let. Rané fáze evoluce pterosauřího letu však zůstávají zcela neznámé, což je dáno především absencí známých blízkých příbuzných (s pravděpodobnou výjimkou skleromochla) a "přechodných forem". Slibný materiál sice existuje v podobě taxonů "Pteromimus" a "Procoelosaurus" z texaského pozdního triasu, mimo předběžného popisu ve formálně nepublikované PhD dizertaci (Atanassov 2002) o nich ale nejsou dostupná žádná další data. Tokita (2014) podotýká, že podobně špatně známé jsou i počátky netopýrů, zároveň ale připomíná, že u této savčí vývojové větve aspoň panuje téměř všeobecná shoda na tom, jak a v jakém prostředí získali svou schopnost letu.
*Faxinalipterus, který rovněž pochází z rozhraní karnu a noru a který měl být podle autorů původního popisu nejbazálnějším známým pterosaurem (Bonaparte et al. 2010), nemá podle ostatních autorů žádné zjevné pterosauří znaky a anatomií pažní kosti se naopak od pterosaurů výrazně odlišuje (Dalla Vecchia 2013); stále však může jít o zástupce Ornithodira (Soares et al. 2013). To se shoduje i s komentáři pterosauřího experta Alexandera Kellnera z roku 2010, podle nějž může jít o bazálního pterosauromorfa nebo o raného zástupce pan-Aves, který předcházel oddělení vývojových větvví pterosaurů a dinosaurů.
Tokita (2014) také stručně připomíná základní strukturu pterosauřího křídla, tvořeného dvěma membránami: menším propatagiem, nataženým mezi krkem a prvním prstem a vyztuženým pteroidem, zápěstní kostí neznámého původu; a větším brachiopatagiem, které se upínalo na špičku extrémně protáhlého čtvrtého prstu a jehož přesný rozsah byl po dlouhou dobu kontroverzní. Exempláře s dochovanými obrysy letových membrán a zbytky měkkých tkání ale ukazují, že se konec brachiopatagia upínal na kotník (Elgin et al. 2011). Stejně jako bazální teropodi, i pterosauři ztratili pátý prst. Extrémní prodloužení čtvrtého prstu podle všeho nebylo doprovázeno redukcí prstů I až III ani výrazným prodloužením jiných segmentů přední končetiny: od ramene až k zápěstí byly přední končetiny pterosaurů jen o něco málo delší než zadní končetiny od kyčle ke kotníku, a teprve u pterodaktyloidů došlo k výraznému protažení záprstních kostí.
Je patrné, že osteologicky se pterosauří křídlo velmi liší od křídla ptáků, a totéž platí i pro myologii. Tokita (2014) uvádí, že ačkoli je ptačí letové svalstvo v poměru k celkové tělesné hmotnosti velmi masivní, jeho uspořádání je relativně jednoduché a dominují mu dva hlavní svaly. Prsní sval (m. pectoralis), který křídlo stlačuje dolů, má začátek na spodní části kýlu hrudní kosti a úpon na přední (proximální) polovině pažní kosti; zatímco podklíčkový nebo hluboký prsní sval (m. supracoracoideus), který křídlo zvedá, má začátek na horní části prsního kýlu a na přilehlém těle hrudní kosti, se přes šlachu procházející skrze trioseální kanál (tvořený lopatkou, kostí pažní a kostí krkavčí) upíná na pažní kost seshora. Naproti tomu u pterosaurů začíná podklíčkový sval na přední straně krkavčí kosti a upíná se na pažní kost přes deltopektorální hřeben, aniž by opsal opsal oblouk okolo glenoidu (jamky ramenního kloubu). Zdá se tedy, že u pterosaurů podklíčkový sval křídlo nezdvihal, nýbrž naopak stlačoval dolů a natáčel. To se shoduje s rolí tohoto svalu u žijících neptačích sauropsidů ("plazů"), a zdá se proto, že svou úlohu v elevaci křídla hluboký prsní sval získal až u ptáků.

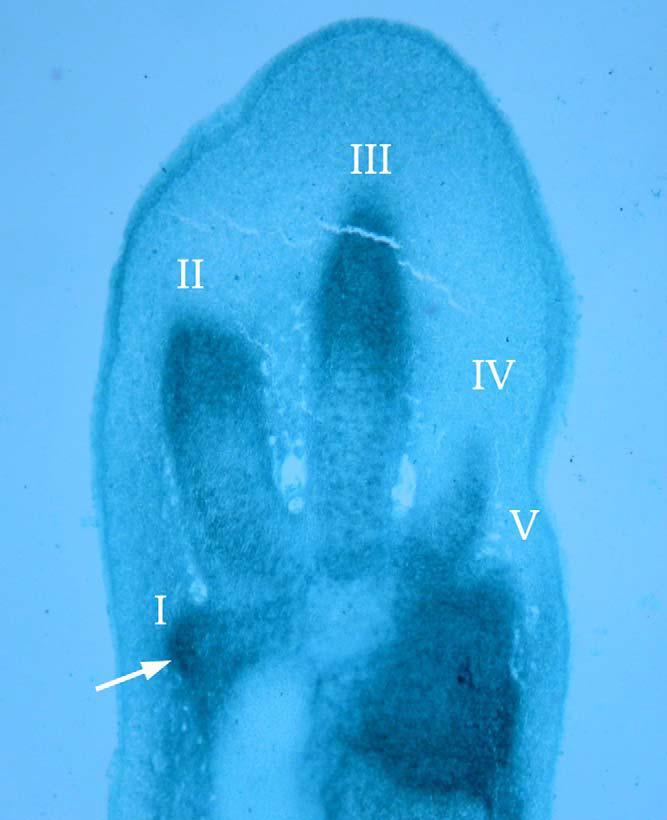
Podle Tokity (2014) lze na základě evolučně-developmentálních dat získaných od žijících tetrapodů přibližně určit, jaké molekulární a buněčné mechanizmy stály za evolucí jedinečné morfologie pterosauřího křídla. Víme, že při formování identity prstů je klíčová tzv. zóna polarizační aktivity (ZPA), oblast mezenchymu na okraji končetinového pupene, a gen Shh (Sonic hedgehog), který je v ní exprimován. První tři prsty (počínaje palcem) vznikají mimo ZPA a to, jakou identitu zaujmou, závisí na tom, jaké koncentraci Shh (proteinu kódovaného genem Shh), difundujícího ze ZPA, budou vystaveny: nejnižší koncentrace specifikují prst I (palec), nejvyšší koncentrace prst III (prostředník). Prsty IV a V vznikají z potomstva buněk uvnitř ZPA, a jejich diferenciaci podle všeho ovlivňuje to, jak dlouho byly vystaveny Shh. (Více jsem o diferenciaci prstů napsal na blogu např. zde). Navzdory tomu, že žádný známý pterosaur pátý prst nemá, je podle Tokity (2014) pravděpodobné, že byl u embrya přítomný jako mezenchymální zárodečná kondenzace: podobně je tomu u žijících ptáků (kde ale číslování kondenzací značně komplikuje debata "axis shift/frame shift": viz Towers et al. 2011; Salinas-Saavedra et al. 2014). K jeho zániku zřejmě docházelo podobným mechanizmem, jako u ptáků.
Zajímavějším problémem je developmentální původ mimořádného prodloužení čtvrtého prstu. Zde se Tokita (2014) obrací k žijícím netopýrům, jejichž křídlo sestává z letové membrány napjaté mezi silně protaženými prsty II až V. Oproti situaci u normálních, krátkoprstých savců dochází ve vyvíjející se ruce netopýrů k řadě významných změn: ZPA je ve stadiu končetinových pupenů (44 dpc – dní po kopulaci; Cretekos et al. 2005; Cooper et al. 2012) prodloužená směrem dopředu (anteriorně); v pozdějším stadiu (50 dpc) dochází ke znovuobnovení exprese Shh, které jinde nemá obdoby; apikální ektodermální lišta, produkující fibroblastický růstový faktor 8 (Fgf8), je rozšířená ve svislém (dorzoventrálním) směru a v relativně pozdním stadiu (50 dpc) zajišťuje navýšení exprese Fgf8 v meziprstních oblastech. Tato zvýšená exprese možná v meziprstních regionech udržuje zpětnovazební smyčku Shh-Fgf, která umožňuje vznik letové membrány a snad i řídí extrémní protažení kostí zadních čtyř prstů. Zatímco ve fázi mezenchymových kondenzací se prsty netopýrů svou délkou nijak neliší od prstů myši, rozdíly nastávají při zchrupavkovatění (chondrifikaci). V perichondriu – husté pojivové tkáni, která obklopuje růstovou ploténku (growth plate) – prodloužených prstů navíc dochází ke zvýšené expresi kostního morfogenetického proteinu 2 (Bmp2), který se na tomto protažení zřejmě také podílí.
Nedávná transkriptomická analýza (Wang et al. 2014) dále ukazuje, že délku netopýřích prstů ovlivňují i homeoboxové (Hox) geny nacházející se na 5' konci shluku D, především pak geny Hoxd11, Hoxd12 a Hoxd13. Netopýři se v porovnání s myší vyznačují extrémně vysokou a dlouhotrvající expresí těchto genů v těch místech embryonální ruky, ze kterých vznikají prsty II až V. Na základě předchozích studií zkoumajících vzájemné vztahy mezi 5' Hoxd, Bmp2 a Shh Tokita navrhuje, že Shh v dopředu rozšířené zóně polarizační aktivity vyvolává prodlouženou expresi 5' Hoxd genů v zadní části růstové ploténky netopýří ruky, a tyto geny zase indukují pozdější expresi Shh v meziprstních oblastech a Bmp2 v meziprstních oblastech a perichondriu, která vede k prodloužení prstních článků zadních čtyř prstů netopýřího křídla. Co nám ale tento scénář může o pterosaurech?
Podle Tokity (2014) je možné, že buňky v ZPA, ze kterých u amniot nakonec vzniká čtvrtý prst, indukovaly lokálně zvýšenou a dlouhotrvající expresi 5' Hoxd, která zase zvýšila produkci kostního morfogenetického proteinu a dalších molekul, čímž urychlila množení a diferenciaci chondrocytů (buněk chrupavky). Na prodloužení čtvrtého prstu se mohly podílet i další geny. Zvláště vhodnými kandidáty se zdají být homeoboxový gen Prx1, který prodlužuje kosti netopýřího křídla; inzulinu podobný růstový faktor IGF1, který ovlivňuje délku dlouhých kostí tím, že reguluje velikost chondrocytů, a jehož vazebný protein 2 (IGFBP2) při zvýšené produkci zkracuje dlouhé kosti kuřecím embryím; Wnt/β-katenin signalizace, která má rozhodující úlohu při formování kloubů (přičemž poloha kloubů vymezuje relativní délku kostí, které tyto klouby oddělují); a konečně hairy2, jehož cyklická exprese zřejmě určuje, kdy se začnou formovat jednotlivé kosti autopodia (zápěstí, záprstí a prstů). Pokud se u pterosaurů změnila perioda exprese hairy2 nebo načasování exprese Wnt, k protažení čtvrtého prstu mohlo dojít už v raných a/nebo pozdních stadiích embryonálního vývoje. To by představovalo významný rozdíl oproti netopýrům, kde se články prstů podpírajících křídlo prodlužují primárně až během postnatálního vývoje. Pterosauři v každém případě museli být unikátní v tom, že jakýkoli mechanizmus zodpovědný za prodlužování prstních článků musel působit výhradně v místech, ze kterých vzniká čtvrtý prst: od žijících amniot takový regulační mechanizmus neznáme.

Druhá hypotéza, kterou Tokita předkládá, navrhuje, že prodloužení čtvrtého prstu bylo pouhým pasivním důsledkem vzniku křídelní membrány, jejíž růst začínal "pučením" epitelu ze stran trupu nazad od předních končetin. U netopýrů tomu tak do jisté míry je: plagiopatagium (největší křídelní membrána netopýrů, napjatá mezi trupem a pátým prstem) vzniká 46 dpc jako laterálně (do stran) zvětšená epitelová struktura, která se teprve později (50 dpc) a po dalším růstu spojí se zárodečným pupenem přední končetiny. Podle Tokity je z tohoto hlediska významné, že stavba letové membrány pterosaurů se do značné míry podobala té netopýří: jak ukazují např. Frey et al. (2003: Figure 6) nebo Witton (2013: Figure 5.14), šlo o překvapivě komplikovanou strukturu, jejíž nejspodnější vrstvu tvořila síť krevních cév, následovaná velmi tenkou prostřední vrstvičkou svalstva a fascie (svalové povázky) a nakonec svrchní vrstvou aktinofibrilů – kolagenových, keratinových nebo snad chrupavčitých vláken, které křídlo vyztužovaly.
Tato podobnost podporuje hypotézu, že stejně jako u netopýrů i u pterosaurů křídlo během embryonálního vývoje rostlo do stran prostřednictvím interakcí mezi kostní a svalovou tkání. Možná to byla právě tkáň brachiopatagia, a zvláště signalizace pomocí fibroblastického růstového faktoru (Fgf10) zodpovědná za její růst, která u pterosauřích embryí způsobovala protažení prstních článků čtvrtého prstu. Podporu této hypotéze dává i transkriptomická analýza Wangové a spol. (2014), která odhalila, že v zárodečném pupenu netopýřího plagiopatagia dochází ke zvýšené a prodloužené expresi genu Tbx3. Ten hned dvěma způsoby podporuje růst prstních článků: jednak potlačuje diferenciaci osteoblastů (kostních buněk), čímž dává chrupavce víc času na růst před tím, než začne osifikovat; jednak zvyšuje expresi genů pro kostní morfogenetické proteiny, který stimuluje množení a diferenciaci chondrocytů. Je možné, že i u pterosaurů exprese Tbx3 v pupenu brachiopatagia aktivovala BMP signalizaci, která spustila prodlužování přilehlého čtvrtého prstu.
V další části své práce se Tokita (2014) věnuje developmentálním mechanizmům, které daly vzniknout masivnímu křídelnímu svalstvu a jeho kostním korelátům (typu robustního ramenního pletence nebo velké, plátovité hrudní kosti). Výzkumy na myších embryích ukazují, že jak pro kostru, tak i pro svalstvo ramenního pletence je zásadní gen Tbx5, jehož absence vede ke kompletní ztrátě předních končetin, lopatky, klíčních kostí a kosti hrudní, a k omezenému a abnormálnímu vývoji pletencových svalů. Anatomických změn typu zvětšení prsního svalu lze tedy zřejmě docílit změnami v regulaci exprese Tbx5. Data získána od kuřat dále ukazují, že významnou roli mohly sehrát i změny v interakcích mezi buňkami svalové a pojivové tkáně. Jak už ale bylo ukázáno výše, analogie mezi pterosaury a ptáky omezuje fakt, že obě skupiny používaly ke stejným účelům odlišné svaly: u pterosaurů to nebyl podklíčkový sval, nýbrž zřejmě svaly začínající na lopatce a zádech, které zvedaly křídlo. Tyto hluboké svaly vznikají u žijících amniot jiným způsobem, než povrchové svaly typu m. pectoralis a m. supracoracoideus, což naznačuje, že ptáci a pterosauři mohli svůj letový aparát vyvinout analogickými, ale opačnými způsoby: ptáci pozměnili vývoj svých povrchových svalů, aniž by byl zasažen vývoj těch hlubokých, zatímco pterosauři možná modifikovali vývoj hlubokých svalů (jako je podlopatkový sval nebo zdvihač lopatky) nezávisle na těch povrchových. Zde ale situaci komplikují obtíže s rekonstruováním myologie fosilních obratlovců. Tokita (2014) dále konstatuje, že protože pterosauři vykazovali pleziomorfické – nikoli ptákům-podobné – uspořádání svalů ramenního pletence, víc užitečných dat nám můžou poskytnout spíše evo-devo studie bazálních tetrapodů než ptáků.
Na vývoji masivního deltopektorálního hřebene pažní kosti, na který se upínalo pterosauří letové svalstvo, se zřejmě podílel gen Scx. Studie na myších a kuřatech ukazují, že exprese tohoto genu v buňkách šlach zvyšuje expresi genu pro kostní morfogenetický protein 4 (Bmp4). Kostní morfogenetický protein 4 vylučovaný buňkami šlach se potom váže na příslušný receptor na přilehlých chondrocytech, čímž indukuje vznik kostních výstupků, na něž se dané šlachy budou upínat. V tomto místě Tokita připomíná, že pterosauři v pozdních stadiích embryonálního vývoje se již zdají mít plně funkční křídla, a byli tedy zřejmě schopni vzlétnout ihned po vylíhnutí (čemuž se ale zdají odporovat data od taxonu Rhamphorhynchus). Podle něj za zvětšením deltopektorálního hřebene pterosaurů stála delší a navýšená exprese Scx ve šlachových buňkách embryí.

Pokud jde o svaly v letových membránách, ty uvnitř brachiopatagia mohly podle Tokity (2014) vznikat z prekurzorů v přední končetině, přičemž jejich uspořádání by determinoval pupen brachiopatagia složený z pojivových tkání – tento scénář je opět založený na analogii s netopýry. Tokita dále poukazuje na fakt, že Wellnhofer (1975) nahlásil zbytky měkké tkáně podobné svalstvu i v menší, přední membráně (propatagiu), a že tyto zbytky připomínají netopýří m. occipito-pollicalis (česky tedy zřejmě "sval týlopalcový"). V případě, že je tato interpretace správná, existovala by pro původ tohoto svalu u pterosaurů hned dvě možná vysvětlení. Případný propatagiální sval mohl vznikat ze stejného prekurzoru, který dává vzniknout zadnímu bříšku dvojbříškatého svalu (m. digastricus posterior) a bodcojazylkovému svalu (m. stylohyoideus) u savců a stahovači dolní čelisti (m. depressor mandibulae) u všech ostatních čelistnatců včetně ptáků*; nebo z prekurzoru zdvihače hlavy (m. sternocleidomastoideus) a trapézového svalu (m. trapezius). V tomto místě se ale zároveň hodí zdůraznit, že přítomnost svalů v pterosauřím propatagiu je poněkud pochybná. Frey et al. (2006), které Tokita necituje, sice skutečně zmínili možnost, že se na špičku pterosauřího pteroidu upínal sval analogický týlopalcovému svalu letounů, Palmer & Dyke (2010) podotýkají, že pterosauří kosti žádné známky po úponech svalů uložených v propatagiu nenesou.
*Přestože ptáci mají sval nazývaný m. stylohyoideus (viz např. Vanden Berge & Zweers 1993), jak konstatovala Tomlinson (2000), tento název je z principu nesprávný, protože ptáci na rozdíl od savců postrádají bodcovitý výběžek (processus styloideus temporalis), po němž je sval pojmenován. Johnston (2011) jej pokládá za homolog m. interhyoideus bazálnějších čelistnatců; Kesteven (1945: 274) jej dokonce jako m. interhyoideus přímo označuje.
K poslední z morfologických změn, jejichž původ se Tokita (2014) snaží odhalit, došlo až uvnitř pterosaurů: jde o výrazné prodloužení záprstních kostí (metakarpálů), které zůstává zřejmě nejlepším diagnostickým znakem skupiny Pterodactyloidea. (Padian [2004] dokonce přímo definoval jméno Pterodactyloidea jako klad vznikající prvním pterosaurem, u kterého bylo záprstí alespoň z 80% tak dlouhé jako pažní kost.) Zde je k dispozici příliš málo dat na navržení konkrétního mechanizmu na molekulární nebo buněčné úrovni, Tokita však alespoň cituje studie, které ukazují, že prstní články a záprstí představují developmentálně samostatné jednotky. Díky tomu mohly procházet modulární evolucí, která zřejmě zodpovídá za protažení záprstních kostí u pterodaktyloidů. Podle Tokity mohlo být v tomto případě klíčovou změnou urychlené množení prekurzorů chrupavkových buněk ve vyvíjejícím se záprstí nebo zrychlení růstu osifikujících metakarpálů.
V závěru své studie autor připomíná, že pterosauři vykazují i řadu jiných znaků, na které by stálo za to se podívat z evo-devo hlediska. Jde např. o ztrátu zubů a souběžný vznik rohovité ramfotéky – Tokita (2014) v tomto bodě cituje studii Loucharta a Viriota (2011), která developmentální bázi těchto evolučních novinek zkoumala u ptáků – nebo známé "pycnofibers", vlákna tvořící pterosauří tělesný pokryv, která mohou a nemusejí být homologická s peřím teropodům a ptakopánvých. Zde autor zmiňuje jako dobrý výchozí bod pro další výzkum rozsáhlou revizi evo-devo ptačího peří od Dhouailly (2009), na blogu už krátce zmiňovanou. Tokita konečně připomíná i již dosažené úspěchy, jako je např. odhad velikosti pterosauřího genomu (Organ & Shedlock 2009), a zdůrazňuje nutnost kombinovat paleontologické poznatky s daty získanými od žijících zvířat, která se s dnešním sekvenováním kompletních genomů rozrůstají dosud nevídaným tempem.
Zdroje:
*Faxinalipterus, který rovněž pochází z rozhraní karnu a noru a který měl být podle autorů původního popisu nejbazálnějším známým pterosaurem (Bonaparte et al. 2010), nemá podle ostatních autorů žádné zjevné pterosauří znaky a anatomií pažní kosti se naopak od pterosaurů výrazně odlišuje (Dalla Vecchia 2013); stále však může jít o zástupce Ornithodira (Soares et al. 2013). To se shoduje i s komentáři pterosauřího experta Alexandera Kellnera z roku 2010, podle nějž může jít o bazálního pterosauromorfa nebo o raného zástupce pan-Aves, který předcházel oddělení vývojových větvví pterosaurů a dinosaurů.
Tokita (2014) také stručně připomíná základní strukturu pterosauřího křídla, tvořeného dvěma membránami: menším propatagiem, nataženým mezi krkem a prvním prstem a vyztuženým pteroidem, zápěstní kostí neznámého původu; a větším brachiopatagiem, které se upínalo na špičku extrémně protáhlého čtvrtého prstu a jehož přesný rozsah byl po dlouhou dobu kontroverzní. Exempláře s dochovanými obrysy letových membrán a zbytky měkkých tkání ale ukazují, že se konec brachiopatagia upínal na kotník (Elgin et al. 2011). Stejně jako bazální teropodi, i pterosauři ztratili pátý prst. Extrémní prodloužení čtvrtého prstu podle všeho nebylo doprovázeno redukcí prstů I až III ani výrazným prodloužením jiných segmentů přední končetiny: od ramene až k zápěstí byly přední končetiny pterosaurů jen o něco málo delší než zadní končetiny od kyčle ke kotníku, a teprve u pterodaktyloidů došlo k výraznému protažení záprstních kostí.
Je patrné, že osteologicky se pterosauří křídlo velmi liší od křídla ptáků, a totéž platí i pro myologii. Tokita (2014) uvádí, že ačkoli je ptačí letové svalstvo v poměru k celkové tělesné hmotnosti velmi masivní, jeho uspořádání je relativně jednoduché a dominují mu dva hlavní svaly. Prsní sval (m. pectoralis), který křídlo stlačuje dolů, má začátek na spodní části kýlu hrudní kosti a úpon na přední (proximální) polovině pažní kosti; zatímco podklíčkový nebo hluboký prsní sval (m. supracoracoideus), který křídlo zvedá, má začátek na horní části prsního kýlu a na přilehlém těle hrudní kosti, se přes šlachu procházející skrze trioseální kanál (tvořený lopatkou, kostí pažní a kostí krkavčí) upíná na pažní kost seshora. Naproti tomu u pterosaurů začíná podklíčkový sval na přední straně krkavčí kosti a upíná se na pažní kost přes deltopektorální hřeben, aniž by opsal opsal oblouk okolo glenoidu (jamky ramenního kloubu). Zdá se tedy, že u pterosaurů podklíčkový sval křídlo nezdvihal, nýbrž naopak stlačoval dolů a natáčel. To se shoduje s rolí tohoto svalu u žijících neptačích sauropsidů ("plazů"), a zdá se proto, že svou úlohu v elevaci křídla hluboký prsní sval získal až u ptáků.
Myologie křídla a ramenního pletence u pterosaurů, ptáků a letounů při pohledu seshora. (A) Rekonstrukce pterosauřího křídla založená na taxonu Anhanguera. Šlacha natahovače křídelního prstu (ligamentum extensor digiti alae) mohla zajišťovat automatické natažení křídelního prstu během letu a zabraňovat hyperextenzi lokte. (B) Křídlo kukačky rýhozobé (Crotophaga sulcirostris). (C) Křídlo kaloně egyptského (Rousettus aegyptiacus). Svaly zvedající křídlo jsou vyznačeny červeně, svaly stlačující křídlo dolů fialově. Některé z létacích svalů nejsou z tohoto pohledu díky svému prostorovému uspořádání viditelné. (Zdroj: Tokita 2014: Figure 2)
Podle Tokity (2014) lze na základě evolučně-developmentálních dat získaných od žijících tetrapodů přibližně určit, jaké molekulární a buněčné mechanizmy stály za evolucí jedinečné morfologie pterosauřího křídla. Víme, že při formování identity prstů je klíčová tzv. zóna polarizační aktivity (ZPA), oblast mezenchymu na okraji končetinového pupene, a gen Shh (Sonic hedgehog), který je v ní exprimován. První tři prsty (počínaje palcem) vznikají mimo ZPA a to, jakou identitu zaujmou, závisí na tom, jaké koncentraci Shh (proteinu kódovaného genem Shh), difundujícího ze ZPA, budou vystaveny: nejnižší koncentrace specifikují prst I (palec), nejvyšší koncentrace prst III (prostředník). Prsty IV a V vznikají z potomstva buněk uvnitř ZPA, a jejich diferenciaci podle všeho ovlivňuje to, jak dlouho byly vystaveny Shh. (Více jsem o diferenciaci prstů napsal na blogu např. zde). Navzdory tomu, že žádný známý pterosaur pátý prst nemá, je podle Tokity (2014) pravděpodobné, že byl u embrya přítomný jako mezenchymální zárodečná kondenzace: podobně je tomu u žijících ptáků (kde ale číslování kondenzací značně komplikuje debata "axis shift/frame shift": viz Towers et al. 2011; Salinas-Saavedra et al. 2014). K jeho zániku zřejmě docházelo podobným mechanizmem, jako u ptáků.
Zajímavějším problémem je developmentální původ mimořádného prodloužení čtvrtého prstu. Zde se Tokita (2014) obrací k žijícím netopýrům, jejichž křídlo sestává z letové membrány napjaté mezi silně protaženými prsty II až V. Oproti situaci u normálních, krátkoprstých savců dochází ve vyvíjející se ruce netopýrů k řadě významných změn: ZPA je ve stadiu končetinových pupenů (44 dpc – dní po kopulaci; Cretekos et al. 2005; Cooper et al. 2012) prodloužená směrem dopředu (anteriorně); v pozdějším stadiu (50 dpc) dochází ke znovuobnovení exprese Shh, které jinde nemá obdoby; apikální ektodermální lišta, produkující fibroblastický růstový faktor 8 (Fgf8), je rozšířená ve svislém (dorzoventrálním) směru a v relativně pozdním stadiu (50 dpc) zajišťuje navýšení exprese Fgf8 v meziprstních oblastech. Tato zvýšená exprese možná v meziprstních regionech udržuje zpětnovazební smyčku Shh-Fgf, která umožňuje vznik letové membrány a snad i řídí extrémní protažení kostí zadních čtyř prstů. Zatímco ve fázi mezenchymových kondenzací se prsty netopýrů svou délkou nijak neliší od prstů myši, rozdíly nastávají při zchrupavkovatění (chondrifikaci). V perichondriu – husté pojivové tkáni, která obklopuje růstovou ploténku (growth plate) – prodloužených prstů navíc dochází ke zvýšené expresi kostního morfogenetického proteinu 2 (Bmp2), který se na tomto protažení zřejmě také podílí.
Nedávná transkriptomická analýza (Wang et al. 2014) dále ukazuje, že délku netopýřích prstů ovlivňují i homeoboxové (Hox) geny nacházející se na 5' konci shluku D, především pak geny Hoxd11, Hoxd12 a Hoxd13. Netopýři se v porovnání s myší vyznačují extrémně vysokou a dlouhotrvající expresí těchto genů v těch místech embryonální ruky, ze kterých vznikají prsty II až V. Na základě předchozích studií zkoumajících vzájemné vztahy mezi 5' Hoxd, Bmp2 a Shh Tokita navrhuje, že Shh v dopředu rozšířené zóně polarizační aktivity vyvolává prodlouženou expresi 5' Hoxd genů v zadní části růstové ploténky netopýří ruky, a tyto geny zase indukují pozdější expresi Shh v meziprstních oblastech a Bmp2 v meziprstních oblastech a perichondriu, která vede k prodloužení prstních článků zadních čtyř prstů netopýřího křídla. Co nám ale tento scénář může o pterosaurech?
Podle Tokity (2014) je možné, že buňky v ZPA, ze kterých u amniot nakonec vzniká čtvrtý prst, indukovaly lokálně zvýšenou a dlouhotrvající expresi 5' Hoxd, která zase zvýšila produkci kostního morfogenetického proteinu a dalších molekul, čímž urychlila množení a diferenciaci chondrocytů (buněk chrupavky). Na prodloužení čtvrtého prstu se mohly podílet i další geny. Zvláště vhodnými kandidáty se zdají být homeoboxový gen Prx1, který prodlužuje kosti netopýřího křídla; inzulinu podobný růstový faktor IGF1, který ovlivňuje délku dlouhých kostí tím, že reguluje velikost chondrocytů, a jehož vazebný protein 2 (IGFBP2) při zvýšené produkci zkracuje dlouhé kosti kuřecím embryím; Wnt/β-katenin signalizace, která má rozhodující úlohu při formování kloubů (přičemž poloha kloubů vymezuje relativní délku kostí, které tyto klouby oddělují); a konečně hairy2, jehož cyklická exprese zřejmě určuje, kdy se začnou formovat jednotlivé kosti autopodia (zápěstí, záprstí a prstů). Pokud se u pterosaurů změnila perioda exprese hairy2 nebo načasování exprese Wnt, k protažení čtvrtého prstu mohlo dojít už v raných a/nebo pozdních stadiích embryonálního vývoje. To by představovalo významný rozdíl oproti netopýrům, kde se články prstů podpírajících křídlo prodlužují primárně až během postnatálního vývoje. Pterosauři v každém případě museli být unikátní v tom, že jakýkoli mechanizmus zodpovědný za prodlužování prstních článků musel působit výhradně v místech, ze kterých vzniká čtvrtý prst: od žijících amniot takový regulační mechanizmus neznáme.
Myologie křídla a ramenního pletence u pterosaurů, ptáků a letounů při pohledu zespoda. (A) Rekonstrukce pterosauřího křídla založená na taxonu Anhanguera. Šlacha natahovače křídelního prstu je z obrázku vynechána. (B) Křídlo kukačky rýhozobé (Crotophaga sulcirostris). (C) Křídlo kaloně egyptského (Rousettus aegyptiacus). Svaly zvedající křídlo jsou vyznačeny červeně, svaly stlačující křídlo dolů fialově. Některé z létacích svalů nejsou z tohoto pohledu díky svému prostorovému uspořádání viditelné. (Zdroj: Tokita 2014: Figure 3)
Tato podobnost podporuje hypotézu, že stejně jako u netopýrů i u pterosaurů křídlo během embryonálního vývoje rostlo do stran prostřednictvím interakcí mezi kostní a svalovou tkání. Možná to byla právě tkáň brachiopatagia, a zvláště signalizace pomocí fibroblastického růstového faktoru (Fgf10) zodpovědná za její růst, která u pterosauřích embryí způsobovala protažení prstních článků čtvrtého prstu. Podporu této hypotéze dává i transkriptomická analýza Wangové a spol. (2014), která odhalila, že v zárodečném pupenu netopýřího plagiopatagia dochází ke zvýšené a prodloužené expresi genu Tbx3. Ten hned dvěma způsoby podporuje růst prstních článků: jednak potlačuje diferenciaci osteoblastů (kostních buněk), čímž dává chrupavce víc času na růst před tím, než začne osifikovat; jednak zvyšuje expresi genů pro kostní morfogenetické proteiny, který stimuluje množení a diferenciaci chondrocytů. Je možné, že i u pterosaurů exprese Tbx3 v pupenu brachiopatagia aktivovala BMP signalizaci, která spustila prodlužování přilehlého čtvrtého prstu.
V další části své práce se Tokita (2014) věnuje developmentálním mechanizmům, které daly vzniknout masivnímu křídelnímu svalstvu a jeho kostním korelátům (typu robustního ramenního pletence nebo velké, plátovité hrudní kosti). Výzkumy na myších embryích ukazují, že jak pro kostru, tak i pro svalstvo ramenního pletence je zásadní gen Tbx5, jehož absence vede ke kompletní ztrátě předních končetin, lopatky, klíčních kostí a kosti hrudní, a k omezenému a abnormálnímu vývoji pletencových svalů. Anatomických změn typu zvětšení prsního svalu lze tedy zřejmě docílit změnami v regulaci exprese Tbx5. Data získána od kuřat dále ukazují, že významnou roli mohly sehrát i změny v interakcích mezi buňkami svalové a pojivové tkáně. Jak už ale bylo ukázáno výše, analogie mezi pterosaury a ptáky omezuje fakt, že obě skupiny používaly ke stejným účelům odlišné svaly: u pterosaurů to nebyl podklíčkový sval, nýbrž zřejmě svaly začínající na lopatce a zádech, které zvedaly křídlo. Tyto hluboké svaly vznikají u žijících amniot jiným způsobem, než povrchové svaly typu m. pectoralis a m. supracoracoideus, což naznačuje, že ptáci a pterosauři mohli svůj letový aparát vyvinout analogickými, ale opačnými způsoby: ptáci pozměnili vývoj svých povrchových svalů, aniž by byl zasažen vývoj těch hlubokých, zatímco pterosauři možná modifikovali vývoj hlubokých svalů (jako je podlopatkový sval nebo zdvihač lopatky) nezávisle na těch povrchových. Zde ale situaci komplikují obtíže s rekonstruováním myologie fosilních obratlovců. Tokita (2014) dále konstatuje, že protože pterosauři vykazovali pleziomorfické – nikoli ptákům-podobné – uspořádání svalů ramenního pletence, víc užitečných dat nám můžou poskytnout spíše evo-devo studie bazálních tetrapodů než ptáků.
Na vývoji masivního deltopektorálního hřebene pažní kosti, na který se upínalo pterosauří letové svalstvo, se zřejmě podílel gen Scx. Studie na myších a kuřatech ukazují, že exprese tohoto genu v buňkách šlach zvyšuje expresi genu pro kostní morfogenetický protein 4 (Bmp4). Kostní morfogenetický protein 4 vylučovaný buňkami šlach se potom váže na příslušný receptor na přilehlých chondrocytech, čímž indukuje vznik kostních výstupků, na něž se dané šlachy budou upínat. V tomto místě Tokita připomíná, že pterosauři v pozdních stadiích embryonálního vývoje se již zdají mít plně funkční křídla, a byli tedy zřejmě schopni vzlétnout ihned po vylíhnutí (čemuž se ale zdají odporovat data od taxonu Rhamphorhynchus). Podle něj za zvětšením deltopektorálního hřebene pterosaurů stála delší a navýšená exprese Scx ve šlachových buňkách embryí.
Evo-devo scénář pro vznik pterosauřího křídla a srovnání s žijícími amnioty. Horní řada: končetina je ve stádiu zárodečného pupene, na jejím okraji se nachází pět zárodečných kondenzací budoucích prstů (šedá kolečka označená P1–5). Kondenzace 4 a 5 se nacházejí v zóně polarizační aktivity (ZPA) vyznačené modře. Vytečkovaná kolečka symbolizují kondenzace, ze kterých žádný prst nevznikne. Druhá řada odshora: prsty nabývají konkrétních morfologických identit. Prsty 4 a 5 vznikají z potomků buněk nacházejících se uvnitř ZPA, které exprimují gen Shh. Prsty 2 a 3 jsou specifikovány koncentrací proteinu Shh difundujícího ze ZPA. Prst 1 vzniká bez účasti Shh. V přední končetině kuřete (třetí sloupek zleva) kondenzace 4 opouští ZPA a přebírá identitu prstu 3 (Tokita se zde přiklání k hypotéze homeotického posunu, contra Towers et al. 2011). V přední končetině netopýra se ZPA rozšiřuje dopředu a nazad od končetinového pupene začíná vznikat zárodek plagiopatagia (krémově), ve kterém dochází k expresi genů Fgf10 a Tbx3. V přední končetině pterosaura zaniká kondenzace 5 a podobně jako u netopýra se nazad od ní objevuje zárodek brachiopatagia (krémově), potenciálně rovněž exprimující Fgf10 a Tbx3. Druhá řada odzdola: boční pohled na embryo v pozdním stadiu vývoje. Dolní řada: detail na končetinu. U netopýra vzniká plagiopatagium (krémově) táhnoucí se od prstu 5 k zadní končetině. Meziprsty nezanikají; místo toho se z nich vyvíjí chiropatagium (růžově), ve kterém dochází k expresi Shh, Hoxd11–13, Fgf8 a Tbx3. Vývoj chiropatagia může současně usnadňovat protažení prstů 2 až 5 (červené šipky). U pterosaura vzniká brachiopatagium (krémově) táhnoucí se od prstu 4 k zadní končetině. Vývoj této membrány může současně usnadňovat protažení prstu 4. (Zdroj: Tokita 2014: Figure 4)
Pokud jde o svaly v letových membránách, ty uvnitř brachiopatagia mohly podle Tokity (2014) vznikat z prekurzorů v přední končetině, přičemž jejich uspořádání by determinoval pupen brachiopatagia složený z pojivových tkání – tento scénář je opět založený na analogii s netopýry. Tokita dále poukazuje na fakt, že Wellnhofer (1975) nahlásil zbytky měkké tkáně podobné svalstvu i v menší, přední membráně (propatagiu), a že tyto zbytky připomínají netopýří m. occipito-pollicalis (česky tedy zřejmě "sval týlopalcový"). V případě, že je tato interpretace správná, existovala by pro původ tohoto svalu u pterosaurů hned dvě možná vysvětlení. Případný propatagiální sval mohl vznikat ze stejného prekurzoru, který dává vzniknout zadnímu bříšku dvojbříškatého svalu (m. digastricus posterior) a bodcojazylkovému svalu (m. stylohyoideus) u savců a stahovači dolní čelisti (m. depressor mandibulae) u všech ostatních čelistnatců včetně ptáků*; nebo z prekurzoru zdvihače hlavy (m. sternocleidomastoideus) a trapézového svalu (m. trapezius). V tomto místě se ale zároveň hodí zdůraznit, že přítomnost svalů v pterosauřím propatagiu je poněkud pochybná. Frey et al. (2006), které Tokita necituje, sice skutečně zmínili možnost, že se na špičku pterosauřího pteroidu upínal sval analogický týlopalcovému svalu letounů, Palmer & Dyke (2010) podotýkají, že pterosauří kosti žádné známky po úponech svalů uložených v propatagiu nenesou.
*Přestože ptáci mají sval nazývaný m. stylohyoideus (viz např. Vanden Berge & Zweers 1993), jak konstatovala Tomlinson (2000), tento název je z principu nesprávný, protože ptáci na rozdíl od savců postrádají bodcovitý výběžek (processus styloideus temporalis), po němž je sval pojmenován. Johnston (2011) jej pokládá za homolog m. interhyoideus bazálnějších čelistnatců; Kesteven (1945: 274) jej dokonce jako m. interhyoideus přímo označuje.
K poslední z morfologických změn, jejichž původ se Tokita (2014) snaží odhalit, došlo až uvnitř pterosaurů: jde o výrazné prodloužení záprstních kostí (metakarpálů), které zůstává zřejmě nejlepším diagnostickým znakem skupiny Pterodactyloidea. (Padian [2004] dokonce přímo definoval jméno Pterodactyloidea jako klad vznikající prvním pterosaurem, u kterého bylo záprstí alespoň z 80% tak dlouhé jako pažní kost.) Zde je k dispozici příliš málo dat na navržení konkrétního mechanizmu na molekulární nebo buněčné úrovni, Tokita však alespoň cituje studie, které ukazují, že prstní články a záprstí představují developmentálně samostatné jednotky. Díky tomu mohly procházet modulární evolucí, která zřejmě zodpovídá za protažení záprstních kostí u pterodaktyloidů. Podle Tokity mohlo být v tomto případě klíčovou změnou urychlené množení prekurzorů chrupavkových buněk ve vyvíjejícím se záprstí nebo zrychlení růstu osifikujících metakarpálů.
V závěru své studie autor připomíná, že pterosauři vykazují i řadu jiných znaků, na které by stálo za to se podívat z evo-devo hlediska. Jde např. o ztrátu zubů a souběžný vznik rohovité ramfotéky – Tokita (2014) v tomto bodě cituje studii Loucharta a Viriota (2011), která developmentální bázi těchto evolučních novinek zkoumala u ptáků – nebo známé "pycnofibers", vlákna tvořící pterosauří tělesný pokryv, která mohou a nemusejí být homologická s peřím teropodům a ptakopánvých. Zde autor zmiňuje jako dobrý výchozí bod pro další výzkum rozsáhlou revizi evo-devo ptačího peří od Dhouailly (2009), na blogu už krátce zmiňovanou. Tokita konečně připomíná i již dosažené úspěchy, jako je např. odhad velikosti pterosauřího genomu (Organ & Shedlock 2009), a zdůrazňuje nutnost kombinovat paleontologické poznatky s daty získanými od žijících zvířat, která se s dnešním sekvenováním kompletních genomů rozrůstají dosud nevídaným tempem.
Zdroje:
- http://ipevs.org.br/blog/?p=1694
- http://scienceblogs.com.br...2010/08/faxinalipterus-minima-o-reptil-voador-mais-antigo...
- http://embryology.med.unsw.edu.au/embryology/index.php?title=Bat_Development
- Atanassov MN 2002 Two new archosaur reptiles from the Late Triassic of Texas. Texas Tech Univ: Unpubl. PhD Thesis. 352 p
- Bonaparte JF, Schultz CL, Soares MB 2010 Pterosauria from the Late Triassic of southern Brazil. 63–71 in Bandyopadhyay S, ed. New Aspects of Mesozoic Biodiversity, Lecture Notes in Earth Sciences Vol. 132. Berlin/Heidelberg: Springer-Verlag
- Cooper LN, Cretekos CJ, Sears KE 2012 The evolution and development of mammalian flight. Wiley Interdiscip Rev: Dev Biol 1(5): 773–9
- Cretekos CJ, Weatherbee SD, Chen C-H, Badwaik NK, Niswander L, Behringer RR, Rasweiler JJ 2005 Embryonic staging system for the short-tailed fruit bat, Carollia perspicillata, a model organism for the mammalian order Chiroptera, based upon timed pregnancies in captive-bred animals. Dev Dyn 233(3): 721–38
- Dalla Vecchia FM 2013 Triassic pterosaurs. 119–55 in Nesbitt SJ, Desojo JB, Irmis RB, eds. Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin. Geol Soc Lond Spec Pub 379
- Dhouailly D 2009 A new scenario for the evolutionary origin of hair, feather, and avian scales. J Anat 214(4): 587–606
- Dyke GJ, de Kat R, Palmer C, van der Kindere J, Naish D, Ganapathisubramani B 2013 Aerodynamic performance of the feathered dinosaur Microraptor and the evolution of feathered flight. Nature Comms 4: 2489
- Elgin RA, Hone DWE, Frey E 2011 The extent of the pterosaur flight membrane. Acta Palaeont Pol 56(1): 99–111
- Frey E, Buchy M-C, Stinnesbeck W, González-González AH, di Stefano A 2006 Muzquizopteryx coahuilensis n.g., n. sp., a nyctosaurid pterosaur with soft tissue preservation from the Coniacian (Late Cretaceous) of northeast Mexico (Coahuila). Oryctos 6: 19–40
- Frey E, Tischlinger H, Buchy M-C, Martill DM 2003 New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion. 233–66 in Buffetaut E, Mazin J-M, eds. Evolution and Palaeobiology of Pterosaurs. Geol Soc Lond Spec Pub 217
- Johnston P 2011 New morphological evidence supports congruent phylogenies and Gondwana vicariance for palaeognathous birds. Zool J Linn Soc 163(3): 959–82
- Kesteven LH 1945 The evolution of the skull and the cephalic muscles: a comparative study of their development and adult morphology. Part III. The Sauria (continued). The Aves. Austral Mus Mem 8(4): 270–93
- Liu Y-Q, Kuang H-W, Jiang X-J, Peng N, Xu H, Sun H-Y 2012 Timing of the earliest known feathered dinosaurs and transitional pterosaurs older than the Jehol Biota. Palaeogeogr Palaeoclimatol Palaeoecol 323–5: 1–12
- Longrich NR, Vinther J, Meng Q-J, Li Q-G, Russell AP 2012 Primitive wing feather arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Curr Biol 22(23): 2262–7
- Louchart A, Viriot L 2011 From snout to beak: the loss of teeth in birds. Trends Ecol Evol 26(12): 663–73
- Organ CL, Shedlock AM 2009 Palaeogenomics of pterosaurs and the evolution of small genome size in flying vertebrates. Biol Lett 5(1): 47–50
- Padian K 2004 The nomenclature of Pterosauria (Reptilia, Archosauria). 27 in Laurin M, ed. Abstracts, First International Phylogenetic Nomenclature Meeting, Paris, Muséum National d’Histoire Naturelle, July 6–9, 2004. Paris: Mus Natl d’Hist Nat
- Palmer C, Dyke GJ 2010 Biomechanics of the unique pterosaur pteroid. Proc R Soc B 277(1684): 1121–7
- Salinas-Saavedra M, Gonzalez-Cabrera C, Ossa-Fuentes L, Botelho JF, Ruiz-Flores M, Vargas AO 2014 New developmental evidence supports a homeotic frameshift of digit identity in the evolution of the bird wing. Front Zool 11: 33
- Soares MB, Dalla Vecchia FM, Schultz CL, Kellner AWA 2013 On the supposed pterosaurian nature of Faxinalipterus minima Bonaparte et al. (2010) from the Upper Triassic of Rio Grande do Sul, Brazil. 95–7 in Sayão JM, Costa FR, Bantim RAM, Kellner AWA, eds. Rio Ptero 2013 – International Symposium on Pterosaurs, Short Communications. Rio de Janeiro: Univ Fed Rio de Janeiro, Mus Nacional
- Sorkin B 2014 Aerial ability in basal Deinonychosauria. Bull Gunma Mus Nat Hist 18: 21–34
- Tomlinson CAB 2000 Feeding in paleognathous birds. 359–94 in Schwenk K, ed. Feeding: Form, Function, and Evolution in Tetrapod Vertebrates. San Diego: Acad Press
- Tokita M 2014 How the pterosaur got its wings. Biol Rev doi:10.1111/brv.12150
- Towers M, Signolet J, Sherman A, Sang H, Tickle C 2011 Insights into bird wing evolution and digit specification from polarizing region fate maps. Nature Comms 2: 426
- Upchurch P, Andres B, Butler RJ, Barrett PM 2014 An analysis of pterosaurian biogeography: implicationsfor the evolutionary history and fossil record quality ofthe first flying vertebrates. Hist Biol doi:10.1080/08912963.2014.939077
- Vanden Berge JC, Zweers GA 1993 Myologia. 189–250 in Baumel JJ, King AS, Breazile JE, Evans HE, Vanden Berge JC, eds. Handbook of Avian Anatomy: Nomina Anatomica Avium, 2nd ed. Cambridge, MA: Pub Nuttall Orthith Club 23
- Wang Z, Dai M-Y, Wang Y, Cooper KL, Zhu T-T, Dong D, Zhang J-P, Zhang S-Y 2014 Unique expression patterns of multiple key genes associated with the evolution of mammalian flight. Proc R Soc B 281(1783): 20133133
- Wellnhofer P 1975 Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. I: Allgemeine Skelettmorphologie. Palaeontogr 148: 1–33
- Witton WP 2013 Pterosaurs: Natural History, Evolution, Anatomy. Princeton: Princeton Univ Press