
1. Dlouhý, táhlý úvod
Začněme zeširoka.
Začněme zeširoka.
Přestože má hypotéza, podle níž jsou ptáci malí, agilní teropodní dinosauři, kteří přežili katastrofu na přelomu křídy a paleogénu, ohromnou vysvětlovací schopnost a od jejího vzkříšení v 70. letech minulého století ji podpořilo ohromné množství důkazů, setkala se s několika tvrdými oříšky. Některé z nich byly vyřešeny tak, jak se paleontologické problémy vyřeší většinou (novými fosiliemi), o jednom se ale vedou rozsáhlé diskuze dodnes. Mám na mysli slavný "problém homologie prstů".
Na křídlech skoro všech ptáků najdeme tři prsty. Jelikož je však ptačí ruka vysoce odvozená (čili divná), bylo překvapivě komplikované určit, které prsty to vlastně jsou. Až v 50. letech 20. století se došlo k závěru, že jde o prsty 2, 3, 4, čili o homology lidského ukazováku, prostředníku a prsteníku. Ovšem už u těch nejranějších dinosaurů, jako jsou Eoraptor a Herrerasaurus, je jasně patrná redukce čtvrtého a pátého prstu: z pátého zbyla v podstatě obnažená záprstní kost bez prstních článků a čtvrtý prst si udržel prstní článek jeden, dosti rudimentární (Langer 2004). Čtvrtý prst se v teropodní evoluci ještě nějakou dobu udržel, a jeho poslední zbytek – holý metakarpál, neboli záprstní kost – zmizel u samé báze kladu Coelurosauria (který zahrnuje ptáky, nejrůznější "raptory", lenochodovité terizinosaury, pštrosovité dinosaury a tyrannosauroidy). To neponechává příliš prostoru nejasnostem: jsou-li ptáci odvození teropodi, pak jejich tři prsty musí být I, II, III. Embryologie a paleontologie tady, zdá se, dospěly k neřešitelnému konfliktu, na němž si neváhali přihřát polívčičku notoričtí popírači dinosauřího původu ptáků. Jejich alternativa, tj. vývoj z neurčitých triasových archosauromorfů, takovým problémem netrpí; i přes chronickou neschopnost BANDitů říct cokoli bližšího o vlastních kandidátech na ptačí předky je totiž jasné, že zvířata to byla bezesporu primitivně pětiprstá.
Otázka, jak se vůbec zárodečné kondenzace povedlo identifikovat, je poměrně komplikovaná a souvisí i s takovými problémy, jako je původ tetrapodů, resp. jak poznat, kterým kostem v ploutvích "svaloploutvých ryb" jsou kosti tetrapodní končetiny homologické. Zatímco obdobu tetrapodní kosti pažní, loketní a vřetenní najdeme i v "rybí" ploutvi velmi snadno, čím víc postupujeme distálně (tedy směrem od trupu), tím víc se věci komplikují. Tam, kde jsou kosti tetrapodů hezky uspořádané do několika pásů (tvořených postupně proximálními a distálními zápěstními kostmi [karpály], záprstními kostmi [metakarpály] a nakonec samotnými prsty), mají "ryby" desítky kostiček, které jako by se odvětvovaly z nějaké centrální osy procházející podélně ploutví. Pátralo se tedy po tom, kudy tato centrální osa, zvaná metapterygiální, prochází v tetrapodní končetině. Vybíhá vůbec za úroveň loketní a vřetenní kosti? Pokud ano, které kosti na ní leží?
Holmgren – mimochodem jeden z prvních autorů, který ptačí prsty identifikoval jako 2, 3, 4 – navrhl, že u tetrapodů se osa v zápěstní oblasti ohýbá a že prsty představují od základu nové (neomorfické) struktury, "přišroubované" k jejímu konci a postrádající jakékoli homology v ploutevních kůstkách "svaloploutvých ryb" (Holmgren 1933). Jak ale ve své revizi celé debaty popisují Wagner & Larsson (2007), časem se uchytil spíš model Watsona (1913), podle nějž metapterygiální osa prochází čtvrtým z pěti tetrapodních prstů a v končetině tetrapodů tedy nenajdeme nic, co by v zásadě nešlo odvodit z ploutevních kůstek ancestrálních "ryb". Tento model byl silně podpořen objevem tzv. primární osy, podél které chondrifikuje (chrupavkovatí) embryonální končetina. Jak ukázali Shubin & Alberch (1986), tato osa prochází kostí pažní a loketní, proximální zápěstní kostí zvanou ulnare a dále v naprosté většině případů čtvrtým distálním karpálem a prstem IV. Neplatí to ovšem pro mloky, kde primární osa prochází prochází prstem II, nebo se spíš – jak podotkli Burke et al. (1998) – v zápěstní oblasti úplně vytrácí. U všech prozkoumaných žab a amniot na ní nicméně leží prst 4, a pokud by tomu tak bylo i u ptáků, znamenalo by to, že dva prsty, které se vzápětí objevují vedle něj, musejí být číslovány jako 2 a 3.

A přesto – když už se primární osa prokazatelně vychýlila jednou, není důvod, aby se nemohla vychýlit znovu. Právě s takovou argumentací přišel vynálezce celého konceptu, Neil Shubin, když se ptačím prstům začal věnovat. Už Shubin a Alberch (1986) si povšimli, že se vznik údajného ptačího prstu 5 (který sice zchrupavkovatí, ale posléze zaniká) radikálně liší od vzniku téhož elementu u všech jiných tetrapodů. Možná tedy vůbec nejde o prst 5, ale o prst 4, což by ovšem znamenalo, že primární osa musí procházet prstem 3. Došlo tedy k jejímu posunu, a hypotéza proto dostala jméno "axis-shift". Její příznivce jsem už vyjmenoval v jednom z předchozích článků; hlavní představa byla taková, že posun osy způsobila "změna ve vývojovém načasování formace prstů". Tím pádem je v pořádku, že se po straně nejdříve zformovaného prstu objevují dva další (protože jsou to staré dobré teropodní prsty I a II). I evidence by se našla – Chatterjee (1998) a Witmer (2002)* třeba upozornili na to, že ulnare, součást primární osy, v ptačí ruce mizí a nahrazuje jej "element X" neznámého původu, což jaksi svádí k myšlence, že primární osa buď vůbec dál nepokračuje, nebo je celá rozhozená. Jenže kritika byla zdrcující – jednoznačně je výchylka primární osy známá jen od mloků, kde některé autory vedla až k návrhům tak extrémním, jako je difylie tetrapodů a tedy dvojitý přechod "ryb" na souš. (Viz např. Holmgrena [1933] – jednou to měli být bahníci, z nichž se stali mloci, podruhé pak lalokoploutví, kteří dali vznik "eutetrapodům", tj. žábám a amniotům.)
Poté přišli se spásnou myšlenkou Wagner & Gauthier (1999). Podle nich došlo v evoluci ptáků k události zvané homeotický posun (frame shift), jejíž existence logicky vyplývá z pokroků v evolučně-developmentální biologii. Ta ukázala, že "pozice" a "identita" prstu jsou překvapivě nesouvisející záležitosti. Obě závisejí na rozdílných genových interakcích probíhajících v rozdílnou dobu, a dokonce i s našimi omezenými znalostmi není vůbec těžké docílit toho, aby jeden prst vyrostl na místě (= z tzv. "zárodečné kondenzace") prstu druhého. Máme také důkazy o tom, že k homeotickému posunu čas od času v evoluci dojde přirozenou cestou; a ví se dokonce, že k jednomu takovému případu došlo zrovna u ptáků. Citovali jej už Wagner a Gauthier: u některých kiviů vzniká prst I na jiné kondenzaci, než je obvyklé – přijmeme-li na chvíli hypotézu obou autorů, podle níž už u "normálních" ptáků prsty vyrůstají odjinud, než by měly (prst I z kondenzace 2, prst II z kondenzace 3 atd.), pak u těchto mutantních kiviů vyrůstá palec z kondenzace 3. Šlo by tedy trochu zjednodušeně říct, že kiviům občas naroste palec tam, kde má slušně vychovaný tetrapod (jako třeba člověk) prostředník.
V roce 1999 byla taková hypotéza pouze méně parsimonní (úspornou) alternativou axis-shiftu, který se obešel bez nutnosti přesunout prstní identity na jiné kondenzace. Jenže embryologové brzy získali k určení pozice prstů lepší metodu, než je spoléhání se na zakonzervovanou pozici metapterygiální osy. Zatímco dřívější zobrazovací metody založené na radioaktivním síranu 35SO4 nebo alciánové modři dokázaly detekovat pouze mimobuněčnou matrix, která vzniká až zchrupavkovatěním (chondrifikací) zárodečného mezenchymu, modernější postupy takto omezené nejsou a dovedou zachytit tvary mezenchymu ještě před tím, než se jeho buňky začnou shlukovat do "předchrupavkových" kondenzací. A s nimi byla nakonec odhalena dlouho postrádaná, čistě mezenchymová Anlage (zárodečná kondenzace) 1, která velmi rychle mizí ještě před tím, než se začne formovat chrupavka (Kundrát et al. 2002; Larsson & Wagner 2002). Tento objev vůbec nezapadá do hypotézy o posunu primární osy – pokud u ptáků opravdu prochází prstem 3, pak musí být dotyčná kondenzace pozůstatkem prstu nacházejícího se před palcem. Homeotický posun s tím naopak problém nemá, čímž si obě hypotézy svou pozici rázem prohodily.

I tak se samozřejmě homeotický posun neobešel bez rozsáhlých kontroverzí. Witmer (2002), který neměl čas do své práce zakomponovat nová zjištění o pětiprstosti, tuto hypotézu zamítl jako cirkulární a radši se vrátil k posunu primární osy, podle kterého je vlastně v paleontologii celá pravda a embryologové to celou dobu měli špatně. Mnohým embryologům zase přišlo jednodušší předpokládat, že to mají špatně paleontologové (Kundrát et al. 2002; Galis et al. 2003), z čehož se zrodila tzv. hypotéza pyramidové redukce. Podle ní ptákům z pozic 2, 3, 4 rostou prsty II, III, IV, které ovšem morfologií konvergují s bývalými prsty I, II, III, což chudáky paleontology tak zmátlo, že prsty fosilií očíslovali chybně. Nelze říct, že by argumenty obou táborů nebyly znělé**, ale pro homeotický posun se začala hromadit evidence: Vargas & Fallon (2005) ukázali, že u amniot je prst I unikátní tím, že v něm nedochází k expresi genů HoxD-10, HoxD-11 a HoxD-12, a že u prstu, který ptákům vyrůstá z druhé zárodečné kondenzace, je tomu úplně stejně. (Autoři nicméně nedali frame-shiftu jednoznačnou přednost před axis-shiftem, který navrhli zachránit tím, že by se nadbytečná "kondenzace 1" interpretovala spíš jako prepollex než zbytek zakrslého palce. K tomuto návrhu se v článku ještě vrátíme.) Vargas et al. (2008) zjistili v zásadě totéž porovnáním s nejbližšími ptačími žijícími příbuznými (totiž krokodýly) a Tamura et al. (2011) – studie, o které už byla a v tomto článku ještě bude řeč – snad dokonce vůbec poprvé identifikovali mechanizmus zodpovědný za to, proč z kondenzace 2 vyrůstá prst I a ne prst II.
Pozice hypotézy homeotického posunu se tím podstatně upevnila. Dva roky zpátky ale přišli s novou výzvou Xu et al. (2009) při popisu bizarního zástupce kladu Ceratosauria, limusaura. Už jen návrh, že teropod takto bazální by nám mohl říct cokoli podstatného o homeotickém posunu, byl docela odvážný: Wagner a Gauthier tuto událost původně namapovali na kladogram o dost blíž žijícím ptákům. Xu et al. (2009) si ale hlavně model homeotického posunu pro své potřeby dosti pozměnili: interpretovali jej jako pozvolnou událost, která navíc úplně nesetřela známky po původní pozici prstů. Ta je stále patrná v podobnosti, jakou vykazují tři (hlavní) metakarpály tetanur s metakarpály II, III a IV jiných teropodů; naproti tomu prsty, které na ně navazují, se podobají spíše prstům I, II a III. Podle Xua a kolektivu (ačkoli to ve své studii nikde nenapsali takhle natvrdo) předkové tetanur ztratili prst I a prsty II, III a IV zvolna, krůček po krůčku změnily svou morfologii tak, že ve výsledku byly k neodlišení od prstů I, II a III dřívějších teropodů. Takový návrh vyvolal u zastánců klasického frame-shiftu poprask a především evolučně-developmentální biologové poukazovali na to, že takový proces nemá s homeotickým posunem nic společného; takže např. Young et al. (2011) Xuovu hypotézu ztotožnili s dřívějším modelem pyramidové redukce a zacházeli s ní jako s konkurencí, nikoli podporou hypotézy homeotického posunu.
(Celý vývoj událostí jsem zde poměrně slušně pokryl: o limusaurovi a jeho implikacích pro homologie prstů jsem napsal v samých začátcích blogu, stejně jako jsem okomentoval reakci Wagnera a Gauthiera. Více jsem také napsal o studii Younga a kolektivu, která evidenci pro obě hypotézy důkladně porovnala.)
Celkově ale model homeotického posunu vyšel ze sporu posílen; v neposlední řadě evidencí ze strany experimentálních embryologických studií, jako byla ta výše zmíněná Tamurova. Do modelu byly zakomponovány některé z modifikací navržených autory popisu limusaura, takže Young et al. (2011) např. nově uznávají, že k posunu došlo dřív, než Wagner a Gauthier původně předpokládali – spíš mezi celofyzoidy a averostrany (v node-based smyslu, tj. Ceratosauria + Tetanurae) než mezi megalosauroidy a neotetanurami, a že posun doprovázela tzv. zóna developmentální variability, která se manifestovala např. v podivném uspořádání ruky různých ceratosaurů. Samotný posun ale Young et al. (2011) předložili coby nepochybný fakt, a skutečně se zdálo, že s tolika nezávislými zdroji důkazů je těžké jej napadnout.
2. Towers et al. (2011)
A to byl omyl. Právě to nedávno udělali Towers et al. (2011) ve své studii, publikované v Nature Communications (stejné místo, kde se nedávno objevila i retropozonová studie ptačí fylogeneze).Nemám ji naneštěstí k dispozici kompletní, ale zásluhou D. Marjanoviće se rozsáhlé úryvky z ní objevily na DML. [Editace z 2.10.2011: Za zaslání fulltextu studie děkuji Filipovi Tvrdému, článek jsem díky tomu mohl doplnit o obrázky.]
2.1 Nová evidence – návrat axis-shiftu
Towers a spol. se zaměřují na zónu polarizační aktivity (ZPA) ptačího křídla, což je oblast na okraji zárodečného pupene končetiny (limb bud), která se skládá z buněk exprimujících gen pro protein zvaný Sonic hedgehog. (Ano, to je skutečně Ježek Sonic. Jak se stalo, že má figurka z videohry vlastní protein, zůstává jednou z největších záhad biologie.) U mnoha tetrapodů vzniká ze zóny polarizační aktivity vůbec první prst, který se na končetině objeví, a to je – jak bylo uvedeno výše v souvislosti s metapterygiální osou – prst IV. Experimenty Towerse a spol. přímo navazují na to, co dělali s ptačí rukou Tamura et al. (2011). Ti zjistili, že podíl této oblasti na vývoji předních a zadních končetin je úplně odlišný a že zatímco v nohou prst IV skutečně vzniká z buněk ZPA, na křídle se buňky, z nichž se stane čtvrtý prst, ze ZPA oddělí ještě před tím, než se prsty začnou formovat. To je podle Tamury a spol. vystavuje vysokým koncentracím Shh, které je de facto přeměňují v prst III. Zdálo se tak, že vůbec poprvé byl demonstrován přesný mechanizmus, jak dochází k přepnutí kondenzace tak, aby začala vyvíjet jiný prst, který hledali zastánci homeotického posunu.
Jenže právě zde na Tamurovy experimenty navázali Towers et al. (2011). Ti sledovali buňky ze ZPA po podstatně delší dobu, a to tak, že v nich "zapnuli" expresi genu pro zelený fluorescenční protein (GFP). Pod UV zářením potom mohli sledovat, kde se buňky z polarizačního regionu nakonec octnou. Výsledek byl šokující: buňky, z nichž se stane prst s identitou III, se skutečně ze ZPA vydělují, ale ty, které v zóně zůstanou, vůbec žádný prst nevytvoří, a to i přesto, že leží přímo na primární ose. Místo toho se z nich stanou měkké tkáně na vnějším (posteriorním) okraji posledního prstu. Kondenzace 4 tedy nepřevezme identitu III – ona nikdy nevznikne. Potvrzeno je přitom Tamurovo pozorování, že na nohou je vše v pořádku – snad za to může (autoři tuto větu začínají slovy "we speculate") fakt, že v ZPA křídla umírá více buněk. Nově přidáno je ale pozorování, že i v myší ruce pocházejí měkké tkáně na vnějším okraji 3. prstu ze zóny polarizační aktivity. Pozice primární osy tedy není u obratlovců zakonzervovaná a autoři s hrdostí znovuoživují axis-shift, se kterým to šlo od objevu "kondenzace 1" na ptačí ruce z kopce. (S tím, že až dosud byla hypotéza posunu primární osy "přehlížená", jak autoři tvrdí, bych ale asi nesouhlasil; příznivci "frame-shiftu" ji komentovali hned několikrát [Wagner 2005; Young et al. 2011].) Důsledky ale mohou být ještě dalekosáhlejší: jak na DML napsal D. Marjanović, otřesena je celá hypotéza primární osy. Nelze přitom říct, že by nedostala zabrat už dříve (viz např. Cohn et al. 2002).

2.2 Jak se vyrovnat s kondenzací 1, aneb jsou ptáci primitivně šestiprstí?
Právě kondenzace 1 – ta, kterou posun osy nepředvídal, kdežto homeotický posun ano – představuje problém. Towers et al. (2011) jej berou na vědomí, ale nepřikládají mu velkou váhu. Podle nich je pochybné, zda dotyčný výběžek mezenchymu skutečně představuje zárodek prstu, a citují studii, která nalezla ještě jednu posteriorní kondenzaci (Welten et al. 2005). Ta by podle číslování "2, 3, 4" měla odpovídat prstu 6! Welten a spol. sami takovou šílenost nenavrhli a kondenzaci interpretují poměrně nudně (a pro Towerse a kolektiv spíše nepříznivě), totiž jako pisiform. "We saw no evidence for more than five digit primordia in the chick foot", napsali Welten et al. (2005:23).
Studie Weltenové a spol. je ale pro současnou diskuzi významná ještě něčím. Autoři se totiž jako jedni z mála obrátili od posunů a pyramidových redukcí k tomu nejjednoduššímu, ale také nejpodivnějšímu vysvětlení: ptačí předci měli šest, nikoli pět prstů. Jako vždy se povedlo vyhrabat nějakou starou citaci, která to už navrhla (Schestakowa 1927) a podle které si ptáci rudiment onoho 6. prstu uchovali. Vhodným kandidátem by byl nejen již zmiňovaný pisiform, který leží ve správné pozici, ale i element X. Umožňovalo by to skutečně spoustu problémů zahladit: Herrerasaurus a Eoraptor by mohli mít tři hlavní prsty stejné jako ptáci (II, III, IV) a ty redukované vedle nich by byly nově číslovány jako V a VI. Problém spočívá v tom, že 6 prstů je o 1 prst víc, než kolik nám jich současná představa o společném předkovi všech žijících tetrapodů dovolí. U žijících tetrapodů jsou navíc výjimky z pětiprstosti dosti pochybné – nadějí jsou opět mloci, na jejichž nohou často spatřujeme tzv. "postminimus" (česky zřejmě "zámalíček"?), což je ale asi spíš jen přídavná zánártní kost. Podle některých je dokonce postminimus náš starý známý, pisiform (Gillies 1929). Holmgren (1952) pak nejenže akceptoval ztotožnění obou elementů, ale interpretoval je jako zbytky 6. prstu, přičemž prepollex ("předpalec") měl být prstem sedmým. Jakkoli je to nepravděpodobné, zcela vyloučit to nejde, jelikož fosilní příbuzní tetrapodího korunního kladu (= mlok, člověk, jejich poslední společný předek a všichni jeho potomci) skutečně sedmi prsty disponovali.
Podle konzervativní interpretace je však pisiform prostě přídavný proximální karpál a ani element X nepředstavuje zbytek ptačího zámalíčku. Jak ale upozornil David Marjanović ve své reakci na studii Weltenové a spol., to ještě neznamená, že se musíme hypotézy o ptačí šestiprstosti vzdát. Jen místo číslování "1, 2, 3, 4, 5, zámalíček" zkusme raději "předpalec, 1, 2, 3, 4, 5". Prepollex je skutečně znám od celé řady tetrapodů, ptákům ovšem většinou hodně vzdálených: mají ho žáby, vačice, morčata a snad i ještěrka (Steiner 1934). Efemérní anteriorní kondenzaci na ruce ptačího embrya prepollex připomíná svým rychlým zánikem a drobnými rozměry, přestože u žab může osifikovat v několik elementů celkem podobných prstním článkům a i tzv. os falciforme, "falešný palec" krtků, vzniká z prepollexu. Takto interpretovaná "kondenzace 1" nás vrací zpátky k "axis-shiftu". Vysvětluje např. daleko lépe, proč neznáme žádné šestiprsté fosilní archosaury: nemít prepollex je úplně normální, nemít prst I a ponechat si místo toho prst VI je ovšem neslýchané. I záhadný element X by mohl být v novém číslování odvysvětlen, a to jako pozůstatek pátého prstu (tomu se konec konců podobá tím, že se v ruce bere víceméně odnikud, zatímco to, co bývá jako "prst 5" označováno obvykle, se odděluje z ulnare).
Wagner (2005), jeden ze spoluautorů homeotického posunu, ovšem zareagoval na tyto spekulace velmi rychle. Je-li správným výkladem "prepollex, 1, 2, 3, 4, 5" (a osa prochází kondenzací 3), měli bychom být schopní to otestovat na embryích nejbližších žijících ptačích příbuzných, krokodýlů. Pokud u nich s metodami, s nimiž jsme dokázali anteriorní mezenchymální kondenzaci odkrýt u ptáků, najdeme prepollex, který před zchrupavkovatěním z ruky vymizí, šlo by o silnou podporu teorie posunu primární osy. Tento experiment provedli Larsson et al. (2010) a žádnou stopu po předpalci nenašli, což je také jediná námitka, kterou byli schopní proti "axis-shiftu" dát dohromady Young et al. (2011). Towers a spol. se s ní nejsou schopni moc vypořádat. Mají-li skutečně pravdu, musíme předpokládat, že krokodýli prepollex ztratili tak dokonale, že jej nelze pozorovat v žádné fázi vývoje; hlavně jde ale o skvělou motivaci zabývat se celou otázkou dál.
2.3 Od kondenzace k prstu díky ježku Sonicovi
Autoři se rovněž zabývali otázkou, jaké mechanizmy rozhodují u ptačích prstů o výsledné morfologii. Zárodečné kondenzace vznikají na dvou různých místech: uvnitř ZPA (prst 4 a případně prst 5) a mimo něj (prsty 1, 2 a 3, jsou-li přítomny všechny). Podle autorů je toto rozdělení vynucené právě způsobem "přidělování" prstních identit, kterým jsou rozdílné hladiny Sonic hedgehogu. Při parakrinní signalizaci (kdy protein difunduje mezibuněčným prostorem k buňkám v okolí) dovedou buňky mimo ZPA rozeznat jen dostatečně velké rozdíly v koncentraci Shh, a protože buňky v ZPA dovedou proteinu vyprodukovat jen omezené množství, počet takto determinovaných prstních identit je omezen – podle autorů na 3. O tom, jaký prst vznikne z buněk samotné ZPA, rozhoduje signalizace autokrinní, při které buňka působí na vlastní receptory.
Je pochopitelné, že u parakrinní signalizaci zaznamenávají největší koncentraci Shh ty buňky, které jsou ZPA nejblíž, a že koncentrace vzrůstá s tím, jak dlouho necháme buňky příslušný gen exprimovat. Towers et al. (2011) to dokonale ilustrovali sérií experimentů, při nichž si pohráli se zastavováním exprese Shh i růstu zárodečných kondenzací. Při včasném zastavení exprese tak na kuřecí noze dostaneme dva palce – jeden vzniklý ze ZPA a determinovaný autokrinní signalizací, druhý pocházející z buněk mimo ZPA, kterým stejnou identitu přidělila signalizace parakrinní. Při pozdějším zaražení exprese už ZPA stihlo zaujmout identitu prstu II a mimo ní vyrostly dva prsty, z nichž ten odlehlejší je teď palec a ten bližší opět prst II. Ještě později – a máme I, II, III, III, kde ze ZPA pochází druhá "III". Když expresi nezastavíme, vznikne normální kuřecí noha (už se nám po ní stýskalo) I, II, III, IV. Jindy autoři zastavili růst oblasti, z níž normálně vznikají zárodečné kondenzace pro prsty I, II a III, a to tak, aby se zde vyvinul pouze jeden prst. Pak si opět počali hrát s přerušováním exprese Sonic hedgehogu a dostali se postupem časů od dvou palců (I, I; druhé číslo vždy znamená identitu prstu vznikajícího ze ZPA) přes dva ukazováky a prostředníky až ke kombinaci III, IV.
Vše výše uvedené se vztahuje pouze na nohu, která se chová spořádaně. U křídla při zaražení růstu příslušné zóny vznikne ruka s jediným prstem (protože ze ZPA na křídle žádný prst nevzniká), který bude v závislosti na čase přerušení exprese Shh buď I, II nebo III. Podle autorů tato vcelku jednoduchá pravidla určují identitu prstů u všech tetrapodů.

3. Wang et al. (2011)
Wang et al. (2011) mají tak trochu smůlu. Jejich studie, která novým, rafinovaným způsobem dokazuje správnost homeotického posunu, vyšla o nějaké dva měsíce poté, co byl homeotický posun vyvrácen. Tedy pozdě, ale ne dost pozdě, aby mohly být závěry Towerse a spol. ocitovány a případně rozcupovány. Wang et al. (2011) totiž nedokazují homeotický posun jako takový, jen to, že ptačí prsty mají identity I, II, III, což se pod axis-shiftem Towerse a spol. rozumí tak nějak samo sebou – jaké by taky měly mít jiné, když vyrůstají z pozic 1, 2, 3. Přesto si jejich studie zaslouží určitou minimální pozornost. Autoři v ní nasazují postup zvaný transkriptomika, který si na rozdíl od genomiky (zabývající se DNA) a proteomiky (zabývající se veškerými molekulami proteinů, které lze nalézt uvnitř jedné buňky) všímá molekul mRNA, do kterých je genetická informace obsažená v DNA transkribována. Zabývá se tedy především tím, které součástí genomu jsou aktivovány a kdy se tak děje. Autoři tedy vzali prsty z předních a zadních končetin kuřat ve dvou různých stadiích vývoje (5,5 a 7 dní starých), osekvenovali všechny mRNA transkripty, které v nich našli, rozeznali celkem přes 14 000 genů a nad výslednými daty provedli vícerozměrné škálování (MDS). Vynechali všechny geny, jejichž exprese se mezi přední a zadní končetinou lišila, a zjistili, že zbývající geny se shlukují podle dvou kritérií: jednak podle toho, z jak starého kuřete byly sekvenovány, jednak podle toho, zda se nacházejí v palci nebo zbývajících prstech. Ukázalo se přitom, že palec na noze (prst 1/I) a prst, který je palcem z hlediska teorie homeotického posunu (prst 2/I) sdílejí celou řadu vlastností, včetně toho, že v nich nedochází k expresi genů Hoxd12, Hand2, Zic3 a Lhx9, z nichž ty první dva jsou přitom dobře známými negativními markery nejvíce anteriorního prstu (v lidštině: když nejsou exprimovány, je to palec).
Wang et al. (2011) kupodivu vůbec nebyli schopni identifikovat identity dvou zbývajících prstů na křídle. Povedlo se identifikovat několik genů, které spojují prsty II a III na křídle s prsty III a IV na noze (v tomto pořadí), v obou případech jsou však kontradikovány signálem jiných genů, který křídelní prst II spojuje s pozdním prstem II nohy a křídelní prst III s tímtéž prstem na noze u mladšího exempláře. Autoři si také povšimli, že křídelní prst III je jedinečný tím, že v něm dochází k expresi genu Socs2, což souhlasí s tím, co o něm nahlásili Tamura et al. (2011) – že je celý developmentálně divný a že má zřejmě vlastní, odvozenou identitu. Obecně se prsty na křídle zdají být podstatně více diferenciované než prsty na zadní končetině. Právě tato zjištění (a ne "potvrzení" frame-shiftu, které vyplývá už z toho, že se na studii podíleli Young a Wagner) se zdají být na studii Wangové a spol. nejcennější.
*Witmerův článek v některých místech prostě nejde nemilovat:
Zdroje:
Otázka, jak se vůbec zárodečné kondenzace povedlo identifikovat, je poměrně komplikovaná a souvisí i s takovými problémy, jako je původ tetrapodů, resp. jak poznat, kterým kostem v ploutvích "svaloploutvých ryb" jsou kosti tetrapodní končetiny homologické. Zatímco obdobu tetrapodní kosti pažní, loketní a vřetenní najdeme i v "rybí" ploutvi velmi snadno, čím víc postupujeme distálně (tedy směrem od trupu), tím víc se věci komplikují. Tam, kde jsou kosti tetrapodů hezky uspořádané do několika pásů (tvořených postupně proximálními a distálními zápěstními kostmi [karpály], záprstními kostmi [metakarpály] a nakonec samotnými prsty), mají "ryby" desítky kostiček, které jako by se odvětvovaly z nějaké centrální osy procházející podélně ploutví. Pátralo se tedy po tom, kudy tato centrální osa, zvaná metapterygiální, prochází v tetrapodní končetině. Vybíhá vůbec za úroveň loketní a vřetenní kosti? Pokud ano, které kosti na ní leží?
Holmgren – mimochodem jeden z prvních autorů, který ptačí prsty identifikoval jako 2, 3, 4 – navrhl, že u tetrapodů se osa v zápěstní oblasti ohýbá a že prsty představují od základu nové (neomorfické) struktury, "přišroubované" k jejímu konci a postrádající jakékoli homology v ploutevních kůstkách "svaloploutvých ryb" (Holmgren 1933). Jak ale ve své revizi celé debaty popisují Wagner & Larsson (2007), časem se uchytil spíš model Watsona (1913), podle nějž metapterygiální osa prochází čtvrtým z pěti tetrapodních prstů a v končetině tetrapodů tedy nenajdeme nic, co by v zásadě nešlo odvodit z ploutevních kůstek ancestrálních "ryb". Tento model byl silně podpořen objevem tzv. primární osy, podél které chondrifikuje (chrupavkovatí) embryonální končetina. Jak ukázali Shubin & Alberch (1986), tato osa prochází kostí pažní a loketní, proximální zápěstní kostí zvanou ulnare a dále v naprosté většině případů čtvrtým distálním karpálem a prstem IV. Neplatí to ovšem pro mloky, kde primární osa prochází prochází prstem II, nebo se spíš – jak podotkli Burke et al. (1998) – v zápěstní oblasti úplně vytrácí. U všech prozkoumaných žab a amniot na ní nicméně leží prst 4, a pokud by tomu tak bylo i u ptáků, znamenalo by to, že dva prsty, které se vzápětí objevují vedle něj, musejí být číslovány jako 2 a 3.

Metapterygiální osa (červeně) probíhající ploutví australského bahníka Neoceratodus (A) a končetinou primitivně pětiprstého amniota (B), z jehož ruky byly pro zjednodušení vynechány zápěstní kosti. Pokud skutečně probíhá prstem 4, můžeme zbývající prsty snadno identifikovat. (Modifikováno z Cohn et al. 2002: Figure 1 a Towers et al. 2011: Figure 1)
A přesto – když už se primární osa prokazatelně vychýlila jednou, není důvod, aby se nemohla vychýlit znovu. Právě s takovou argumentací přišel vynálezce celého konceptu, Neil Shubin, když se ptačím prstům začal věnovat. Už Shubin a Alberch (1986) si povšimli, že se vznik údajného ptačího prstu 5 (který sice zchrupavkovatí, ale posléze zaniká) radikálně liší od vzniku téhož elementu u všech jiných tetrapodů. Možná tedy vůbec nejde o prst 5, ale o prst 4, což by ovšem znamenalo, že primární osa musí procházet prstem 3. Došlo tedy k jejímu posunu, a hypotéza proto dostala jméno "axis-shift". Její příznivce jsem už vyjmenoval v jednom z předchozích článků; hlavní představa byla taková, že posun osy způsobila "změna ve vývojovém načasování formace prstů". Tím pádem je v pořádku, že se po straně nejdříve zformovaného prstu objevují dva další (protože jsou to staré dobré teropodní prsty I a II). I evidence by se našla – Chatterjee (1998) a Witmer (2002)* třeba upozornili na to, že ulnare, součást primární osy, v ptačí ruce mizí a nahrazuje jej "element X" neznámého původu, což jaksi svádí k myšlence, že primární osa buď vůbec dál nepokračuje, nebo je celá rozhozená. Jenže kritika byla zdrcující – jednoznačně je výchylka primární osy známá jen od mloků, kde některé autory vedla až k návrhům tak extrémním, jako je difylie tetrapodů a tedy dvojitý přechod "ryb" na souš. (Viz např. Holmgrena [1933] – jednou to měli být bahníci, z nichž se stali mloci, podruhé pak lalokoploutví, kteří dali vznik "eutetrapodům", tj. žábám a amniotům.)
Poté přišli se spásnou myšlenkou Wagner & Gauthier (1999). Podle nich došlo v evoluci ptáků k události zvané homeotický posun (frame shift), jejíž existence logicky vyplývá z pokroků v evolučně-developmentální biologii. Ta ukázala, že "pozice" a "identita" prstu jsou překvapivě nesouvisející záležitosti. Obě závisejí na rozdílných genových interakcích probíhajících v rozdílnou dobu, a dokonce i s našimi omezenými znalostmi není vůbec těžké docílit toho, aby jeden prst vyrostl na místě (= z tzv. "zárodečné kondenzace") prstu druhého. Máme také důkazy o tom, že k homeotickému posunu čas od času v evoluci dojde přirozenou cestou; a ví se dokonce, že k jednomu takovému případu došlo zrovna u ptáků. Citovali jej už Wagner a Gauthier: u některých kiviů vzniká prst I na jiné kondenzaci, než je obvyklé – přijmeme-li na chvíli hypotézu obou autorů, podle níž už u "normálních" ptáků prsty vyrůstají odjinud, než by měly (prst I z kondenzace 2, prst II z kondenzace 3 atd.), pak u těchto mutantních kiviů vyrůstá palec z kondenzace 3. Šlo by tedy trochu zjednodušeně říct, že kiviům občas naroste palec tam, kde má slušně vychovaný tetrapod (jako třeba člověk) prostředník.
V roce 1999 byla taková hypotéza pouze méně parsimonní (úspornou) alternativou axis-shiftu, který se obešel bez nutnosti přesunout prstní identity na jiné kondenzace. Jenže embryologové brzy získali k určení pozice prstů lepší metodu, než je spoléhání se na zakonzervovanou pozici metapterygiální osy. Zatímco dřívější zobrazovací metody založené na radioaktivním síranu 35SO4 nebo alciánové modři dokázaly detekovat pouze mimobuněčnou matrix, která vzniká až zchrupavkovatěním (chondrifikací) zárodečného mezenchymu, modernější postupy takto omezené nejsou a dovedou zachytit tvary mezenchymu ještě před tím, než se jeho buňky začnou shlukovat do "předchrupavkových" kondenzací. A s nimi byla nakonec odhalena dlouho postrádaná, čistě mezenchymová Anlage (zárodečná kondenzace) 1, která velmi rychle mizí ještě před tím, než se začne formovat chrupavka (Kundrát et al. 2002; Larsson & Wagner 2002). Tento objev vůbec nezapadá do hypotézy o posunu primární osy – pokud u ptáků opravdu prochází prstem 3, pak musí být dotyčná kondenzace pozůstatkem prstu nacházejícího se před palcem. Homeotický posun s tím naopak problém nemá, čímž si obě hypotézy svou pozici rázem prohodily.
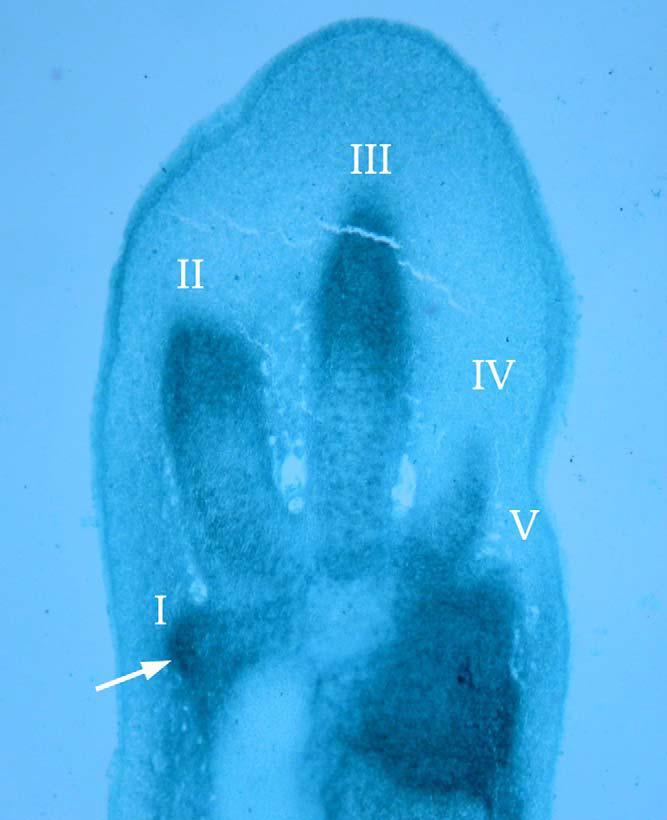
Průřez zárodkem kuřecího křídla. Aplikace moderních embryologických zobrazovacích metod, jako jde zde využitá schopnost aglutininu podzemnice olejné vázat specifické sacharidy obsažené v glykoproteinech na membránách shlukujících se buněk, dovolila odhalit pětiprstý základní stav ptačí ruky. Problém hypotéze o posunu primární osy dělá nejanteriornější kondenzace (vyznačená šipkou a číslovaná zde jako I), která naopak pěkně zapadá do posunu homeotického. Ten ji bere jako důkaz, že ptačí prsty se vyvíjejí z kondenzací 2, 3, 4 (na obrázku zřetelně dominujících). (Zdroj: Wagner 2005: Figure 2)
I tak se samozřejmě homeotický posun neobešel bez rozsáhlých kontroverzí. Witmer (2002), který neměl čas do své práce zakomponovat nová zjištění o pětiprstosti, tuto hypotézu zamítl jako cirkulární a radši se vrátil k posunu primární osy, podle kterého je vlastně v paleontologii celá pravda a embryologové to celou dobu měli špatně. Mnohým embryologům zase přišlo jednodušší předpokládat, že to mají špatně paleontologové (Kundrát et al. 2002; Galis et al. 2003), z čehož se zrodila tzv. hypotéza pyramidové redukce. Podle ní ptákům z pozic 2, 3, 4 rostou prsty II, III, IV, které ovšem morfologií konvergují s bývalými prsty I, II, III, což chudáky paleontology tak zmátlo, že prsty fosilií očíslovali chybně. Nelze říct, že by argumenty obou táborů nebyly znělé**, ale pro homeotický posun se začala hromadit evidence: Vargas & Fallon (2005) ukázali, že u amniot je prst I unikátní tím, že v něm nedochází k expresi genů HoxD-10, HoxD-11 a HoxD-12, a že u prstu, který ptákům vyrůstá z druhé zárodečné kondenzace, je tomu úplně stejně. (Autoři nicméně nedali frame-shiftu jednoznačnou přednost před axis-shiftem, který navrhli zachránit tím, že by se nadbytečná "kondenzace 1" interpretovala spíš jako prepollex než zbytek zakrslého palce. K tomuto návrhu se v článku ještě vrátíme.) Vargas et al. (2008) zjistili v zásadě totéž porovnáním s nejbližšími ptačími žijícími příbuznými (totiž krokodýly) a Tamura et al. (2011) – studie, o které už byla a v tomto článku ještě bude řeč – snad dokonce vůbec poprvé identifikovali mechanizmus zodpovědný za to, proč z kondenzace 2 vyrůstá prst I a ne prst II.
Pozice hypotézy homeotického posunu se tím podstatně upevnila. Dva roky zpátky ale přišli s novou výzvou Xu et al. (2009) při popisu bizarního zástupce kladu Ceratosauria, limusaura. Už jen návrh, že teropod takto bazální by nám mohl říct cokoli podstatného o homeotickém posunu, byl docela odvážný: Wagner a Gauthier tuto událost původně namapovali na kladogram o dost blíž žijícím ptákům. Xu et al. (2009) si ale hlavně model homeotického posunu pro své potřeby dosti pozměnili: interpretovali jej jako pozvolnou událost, která navíc úplně nesetřela známky po původní pozici prstů. Ta je stále patrná v podobnosti, jakou vykazují tři (hlavní) metakarpály tetanur s metakarpály II, III a IV jiných teropodů; naproti tomu prsty, které na ně navazují, se podobají spíše prstům I, II a III. Podle Xua a kolektivu (ačkoli to ve své studii nikde nenapsali takhle natvrdo) předkové tetanur ztratili prst I a prsty II, III a IV zvolna, krůček po krůčku změnily svou morfologii tak, že ve výsledku byly k neodlišení od prstů I, II a III dřívějších teropodů. Takový návrh vyvolal u zastánců klasického frame-shiftu poprask a především evolučně-developmentální biologové poukazovali na to, že takový proces nemá s homeotickým posunem nic společného; takže např. Young et al. (2011) Xuovu hypotézu ztotožnili s dřívějším modelem pyramidové redukce a zacházeli s ní jako s konkurencí, nikoli podporou hypotézy homeotického posunu.
(Celý vývoj událostí jsem zde poměrně slušně pokryl: o limusaurovi a jeho implikacích pro homologie prstů jsem napsal v samých začátcích blogu, stejně jako jsem okomentoval reakci Wagnera a Gauthiera. Více jsem také napsal o studii Younga a kolektivu, která evidenci pro obě hypotézy důkladně porovnala.)
Celkově ale model homeotického posunu vyšel ze sporu posílen; v neposlední řadě evidencí ze strany experimentálních embryologických studií, jako byla ta výše zmíněná Tamurova. Do modelu byly zakomponovány některé z modifikací navržených autory popisu limusaura, takže Young et al. (2011) např. nově uznávají, že k posunu došlo dřív, než Wagner a Gauthier původně předpokládali – spíš mezi celofyzoidy a averostrany (v node-based smyslu, tj. Ceratosauria + Tetanurae) než mezi megalosauroidy a neotetanurami, a že posun doprovázela tzv. zóna developmentální variability, která se manifestovala např. v podivném uspořádání ruky různých ceratosaurů. Samotný posun ale Young et al. (2011) předložili coby nepochybný fakt, a skutečně se zdálo, že s tolika nezávislými zdroji důkazů je těžké jej napadnout.
2. Towers et al. (2011)
A to byl omyl. Právě to nedávno udělali Towers et al. (2011) ve své studii, publikované v Nature Communications (stejné místo, kde se nedávno objevila i retropozonová studie ptačí fylogeneze).
2.1 Nová evidence – návrat axis-shiftu
Towers a spol. se zaměřují na zónu polarizační aktivity (ZPA) ptačího křídla, což je oblast na okraji zárodečného pupene končetiny (limb bud), která se skládá z buněk exprimujících gen pro protein zvaný Sonic hedgehog. (Ano, to je skutečně Ježek Sonic. Jak se stalo, že má figurka z videohry vlastní protein, zůstává jednou z největších záhad biologie.) U mnoha tetrapodů vzniká ze zóny polarizační aktivity vůbec první prst, který se na končetině objeví, a to je – jak bylo uvedeno výše v souvislosti s metapterygiální osou – prst IV. Experimenty Towerse a spol. přímo navazují na to, co dělali s ptačí rukou Tamura et al. (2011). Ti zjistili, že podíl této oblasti na vývoji předních a zadních končetin je úplně odlišný a že zatímco v nohou prst IV skutečně vzniká z buněk ZPA, na křídle se buňky, z nichž se stane čtvrtý prst, ze ZPA oddělí ještě před tím, než se prsty začnou formovat. To je podle Tamury a spol. vystavuje vysokým koncentracím Shh, které je de facto přeměňují v prst III. Zdálo se tak, že vůbec poprvé byl demonstrován přesný mechanizmus, jak dochází k přepnutí kondenzace tak, aby začala vyvíjet jiný prst, který hledali zastánci homeotického posunu.
Jenže právě zde na Tamurovy experimenty navázali Towers et al. (2011). Ti sledovali buňky ze ZPA po podstatně delší dobu, a to tak, že v nich "zapnuli" expresi genu pro zelený fluorescenční protein (GFP). Pod UV zářením potom mohli sledovat, kde se buňky z polarizačního regionu nakonec octnou. Výsledek byl šokující: buňky, z nichž se stane prst s identitou III, se skutečně ze ZPA vydělují, ale ty, které v zóně zůstanou, vůbec žádný prst nevytvoří, a to i přesto, že leží přímo na primární ose. Místo toho se z nich stanou měkké tkáně na vnějším (posteriorním) okraji posledního prstu. Kondenzace 4 tedy nepřevezme identitu III – ona nikdy nevznikne. Potvrzeno je přitom Tamurovo pozorování, že na nohou je vše v pořádku – snad za to může (autoři tuto větu začínají slovy "we speculate") fakt, že v ZPA křídla umírá více buněk. Nově přidáno je ale pozorování, že i v myší ruce pocházejí měkké tkáně na vnějším okraji 3. prstu ze zóny polarizační aktivity. Pozice primární osy tedy není u obratlovců zakonzervovaná a autoři s hrdostí znovuoživují axis-shift, se kterým to šlo od objevu "kondenzace 1" na ptačí ruce z kopce. (S tím, že až dosud byla hypotéza posunu primární osy "přehlížená", jak autoři tvrdí, bych ale asi nesouhlasil; příznivci "frame-shiftu" ji komentovali hned několikrát [Wagner 2005; Young et al. 2011].) Důsledky ale mohou být ještě dalekosáhlejší: jak na DML napsal D. Marjanović, otřesena je celá hypotéza primární osy. Nelze přitom říct, že by nedostala zabrat už dříve (viz např. Cohn et al. 2002).

Pokusy Towerse a kolektivu s kuřecím křídlem. U embryí starých 3 dny, kdy začíná docházet k expresi genu pro Sonic hedgehog, vyměnili původní ZPA za novou, zeleně světélkující. Po 7 se ukázalo, že buňky ze ZPA se nacházejí na vnějším okraji prstu 4 (a). Aby se autoři ujistili, že transplantují skutečně jen ZPA a nic jiného, prohledali dárcovské embryo ihned po odejmutí ZPA na přítomnost transkriptů (molekul mRNA), které indikují, že gen Shh je "zapnutý" a skutečně daný protein produkuje. O 4 hodiny později stejným způsobem analyzovali i embryo, do kterého ZPA přenesli. Ukázalo se, že Shh transkripty v dárcovském embryu zcela chybějí a v příjemci jsou rozeznatelné jen v místě transplantátu (b). Pokud se podíváme na zárodečné křídlo šestidenního embrya seshora, zdá se, že světélkující buňky původem ze ZPA se začínají kondenzovat do chrupavčitého zárodku prstu 4 (c). Podélný (d) a příčný (e) řez příslušnou oblastí desetidenního embrya ale ukáže, že světélkující buňky tvoří pouze měkkou tkáň přilehlou ke chrupavce, nikoli chrupavčitý základ prstu samotný. Obrázek (d) rovněž zachycuje, že buněk exprimujících protein pro fluorescenci ubývá s tím, jak postupujeme směrem od zápěstí ke špičce prstu 4. Autoři také transplantovali světélkující ZPA na vnitřní (anteriorní) stranu končetiny (přirozeně se ZPA vyskytuje jen na straně vnější), čímž zduplikovali prsty do vzorce 4-3-2-2-3-4. ZPA se ale stále podílela pouze na proužku měkkých tkání na vnějším okraji čtvrtého prstu (f, g). V další sérii experimentů autoři transplantovali světélkující ZPA z křídla na vnitřní (h) i vnější (i) stranu nohy, kde – jak je patrné z podélných průřezů (j, k) – dala vzniknout nejen měkkým tkáním, ale také chrupavce, čili kompletnímu prstu. Měřítko: (a, f, i) – 1 mm; (b, c) – 100 μm; (d, g, j, k) – 50 μm; (e) – 150 μm; (h) – 750 μm. (Zdroj: Towers et al. 2011: Figure 2)
2.2 Jak se vyrovnat s kondenzací 1, aneb jsou ptáci primitivně šestiprstí?
Právě kondenzace 1 – ta, kterou posun osy nepředvídal, kdežto homeotický posun ano – představuje problém. Towers et al. (2011) jej berou na vědomí, ale nepřikládají mu velkou váhu. Podle nich je pochybné, zda dotyčný výběžek mezenchymu skutečně představuje zárodek prstu, a citují studii, která nalezla ještě jednu posteriorní kondenzaci (Welten et al. 2005). Ta by podle číslování "2, 3, 4" měla odpovídat prstu 6! Welten a spol. sami takovou šílenost nenavrhli a kondenzaci interpretují poměrně nudně (a pro Towerse a kolektiv spíše nepříznivě), totiž jako pisiform. "We saw no evidence for more than five digit primordia in the chick foot", napsali Welten et al. (2005:23).
Studie Weltenové a spol. je ale pro současnou diskuzi významná ještě něčím. Autoři se totiž jako jedni z mála obrátili od posunů a pyramidových redukcí k tomu nejjednoduššímu, ale také nejpodivnějšímu vysvětlení: ptačí předci měli šest, nikoli pět prstů. Jako vždy se povedlo vyhrabat nějakou starou citaci, která to už navrhla (Schestakowa 1927) a podle které si ptáci rudiment onoho 6. prstu uchovali. Vhodným kandidátem by byl nejen již zmiňovaný pisiform, který leží ve správné pozici, ale i element X. Umožňovalo by to skutečně spoustu problémů zahladit: Herrerasaurus a Eoraptor by mohli mít tři hlavní prsty stejné jako ptáci (II, III, IV) a ty redukované vedle nich by byly nově číslovány jako V a VI. Problém spočívá v tom, že 6 prstů je o 1 prst víc, než kolik nám jich současná představa o společném předkovi všech žijících tetrapodů dovolí. U žijících tetrapodů jsou navíc výjimky z pětiprstosti dosti pochybné – nadějí jsou opět mloci, na jejichž nohou často spatřujeme tzv. "postminimus" (česky zřejmě "zámalíček"?), což je ale asi spíš jen přídavná zánártní kost. Podle některých je dokonce postminimus náš starý známý, pisiform (Gillies 1929). Holmgren (1952) pak nejenže akceptoval ztotožnění obou elementů, ale interpretoval je jako zbytky 6. prstu, přičemž prepollex ("předpalec") měl být prstem sedmým. Jakkoli je to nepravděpodobné, zcela vyloučit to nejde, jelikož fosilní příbuzní tetrapodího korunního kladu (= mlok, člověk, jejich poslední společný předek a všichni jeho potomci) skutečně sedmi prsty disponovali.
Podle konzervativní interpretace je však pisiform prostě přídavný proximální karpál a ani element X nepředstavuje zbytek ptačího zámalíčku. Jak ale upozornil David Marjanović ve své reakci na studii Weltenové a spol., to ještě neznamená, že se musíme hypotézy o ptačí šestiprstosti vzdát. Jen místo číslování "1, 2, 3, 4, 5, zámalíček" zkusme raději "předpalec, 1, 2, 3, 4, 5". Prepollex je skutečně znám od celé řady tetrapodů, ptákům ovšem většinou hodně vzdálených: mají ho žáby, vačice, morčata a snad i ještěrka (Steiner 1934). Efemérní anteriorní kondenzaci na ruce ptačího embrya prepollex připomíná svým rychlým zánikem a drobnými rozměry, přestože u žab může osifikovat v několik elementů celkem podobných prstním článkům a i tzv. os falciforme, "falešný palec" krtků, vzniká z prepollexu. Takto interpretovaná "kondenzace 1" nás vrací zpátky k "axis-shiftu". Vysvětluje např. daleko lépe, proč neznáme žádné šestiprsté fosilní archosaury: nemít prepollex je úplně normální, nemít prst I a ponechat si místo toho prst VI je ovšem neslýchané. I záhadný element X by mohl být v novém číslování odvysvětlen, a to jako pozůstatek pátého prstu (tomu se konec konců podobá tím, že se v ruce bere víceméně odnikud, zatímco to, co bývá jako "prst 5" označováno obvykle, se odděluje z ulnare).
Wagner (2005), jeden ze spoluautorů homeotického posunu, ovšem zareagoval na tyto spekulace velmi rychle. Je-li správným výkladem "prepollex, 1, 2, 3, 4, 5" (a osa prochází kondenzací 3), měli bychom být schopní to otestovat na embryích nejbližších žijících ptačích příbuzných, krokodýlů. Pokud u nich s metodami, s nimiž jsme dokázali anteriorní mezenchymální kondenzaci odkrýt u ptáků, najdeme prepollex, který před zchrupavkovatěním z ruky vymizí, šlo by o silnou podporu teorie posunu primární osy. Tento experiment provedli Larsson et al. (2010) a žádnou stopu po předpalci nenašli, což je také jediná námitka, kterou byli schopní proti "axis-shiftu" dát dohromady Young et al. (2011). Towers a spol. se s ní nejsou schopni moc vypořádat. Mají-li skutečně pravdu, musíme předpokládat, že krokodýli prepollex ztratili tak dokonale, že jej nelze pozorovat v žádné fázi vývoje; hlavně jde ale o skvělou motivaci zabývat se celou otázkou dál.
2.3 Od kondenzace k prstu díky ježku Sonicovi
Autoři se rovněž zabývali otázkou, jaké mechanizmy rozhodují u ptačích prstů o výsledné morfologii. Zárodečné kondenzace vznikají na dvou různých místech: uvnitř ZPA (prst 4 a případně prst 5) a mimo něj (prsty 1, 2 a 3, jsou-li přítomny všechny). Podle autorů je toto rozdělení vynucené právě způsobem "přidělování" prstních identit, kterým jsou rozdílné hladiny Sonic hedgehogu. Při parakrinní signalizaci (kdy protein difunduje mezibuněčným prostorem k buňkám v okolí) dovedou buňky mimo ZPA rozeznat jen dostatečně velké rozdíly v koncentraci Shh, a protože buňky v ZPA dovedou proteinu vyprodukovat jen omezené množství, počet takto determinovaných prstních identit je omezen – podle autorů na 3. O tom, jaký prst vznikne z buněk samotné ZPA, rozhoduje signalizace autokrinní, při které buňka působí na vlastní receptory.
Je pochopitelné, že u parakrinní signalizaci zaznamenávají největší koncentraci Shh ty buňky, které jsou ZPA nejblíž, a že koncentrace vzrůstá s tím, jak dlouho necháme buňky příslušný gen exprimovat. Towers et al. (2011) to dokonale ilustrovali sérií experimentů, při nichž si pohráli se zastavováním exprese Shh i růstu zárodečných kondenzací. Při včasném zastavení exprese tak na kuřecí noze dostaneme dva palce – jeden vzniklý ze ZPA a determinovaný autokrinní signalizací, druhý pocházející z buněk mimo ZPA, kterým stejnou identitu přidělila signalizace parakrinní. Při pozdějším zaražení exprese už ZPA stihlo zaujmout identitu prstu II a mimo ní vyrostly dva prsty, z nichž ten odlehlejší je teď palec a ten bližší opět prst II. Ještě později – a máme I, II, III, III, kde ze ZPA pochází druhá "III". Když expresi nezastavíme, vznikne normální kuřecí noha (už se nám po ní stýskalo) I, II, III, IV. Jindy autoři zastavili růst oblasti, z níž normálně vznikají zárodečné kondenzace pro prsty I, II a III, a to tak, aby se zde vyvinul pouze jeden prst. Pak si opět počali hrát s přerušováním exprese Sonic hedgehogu a dostali se postupem časů od dvou palců (I, I; druhé číslo vždy znamená identitu prstu vznikajícího ze ZPA) přes dva ukazováky a prostředníky až ke kombinaci III, IV.
Vše výše uvedené se vztahuje pouze na nohu, která se chová spořádaně. U křídla při zaražení růstu příslušné zóny vznikne ruka s jediným prstem (protože ze ZPA na křídle žádný prst nevzniká), který bude v závislosti na čase přerušení exprese Shh buď I, II nebo III. Podle autorů tato vcelku jednoduchá pravidla určují identitu prstů u všech tetrapodů.

Názorné vysvětlení rozdílu mezi hypotézami homeotického posunu (frame-shift) a posunu primární osy (axis-shift). Na obrázku (a) zcela nahoře jsou nejprve namapovány ruce různých archosaurů na kladogram (který je trochu divně nakreslený a je nutné jej číst tak, že taxony vpravo jsou si blíž příbuzné než ty, které stojí nalevo od nich). Morfologie prstů – hlavně pak tzv. falangeální formulka, která říká, z kolika prstních článků každý prst sestává – jednoznačně podporují hypotézu, že prsty žijících ptáků jsou I, II, III (charakterizované formulkou 3-4-5). Dole jsou zachyceny rozdíly mezi frame-shiftem a axis-shiftem. Obě hypotézy se shodují na tom, že předci všech amniot a tedy i ptáků (jejichž morfologii dokumentuje výše zachycená Seymouria) měli prsty I, II, III, IV a V vyrůstající z kondenzací 1, 2, 3, 4, 5 (c); přičemž primární osa procházela prstem IV/4 (b). Neshoda panuje na přesném mechanizmu, jímž se z pětiprsté ruky stala ruka tříprstá. Podle homeotického posunu (d, e) se ztratily kondenzace 1 a 5 a prsty I, II, III se přenesly na kondenzace zbývající. Podle posunu osy se ztratily kondenzace 4 a 5, zbývající prsty na svých kondenzacích zůstaly (g), ale primární osa se vychýlila tak, že nyní prochází prstem 3 (f). Tuto hypotézu podporuje fakt, že ze zóny polarizační aktivity nevzniká ve fázích 18 až 21 (tedy u embrya starého 3 až 3,5 dne) prst IV, jak by tomu být mělo. (Zdroj: Towers et al. 2011: Figure 1)
3. Wang et al. (2011)
Wang et al. (2011) mají tak trochu smůlu. Jejich studie, která novým, rafinovaným způsobem dokazuje správnost homeotického posunu, vyšla o nějaké dva měsíce poté, co byl homeotický posun vyvrácen. Tedy pozdě, ale ne dost pozdě, aby mohly být závěry Towerse a spol. ocitovány a případně rozcupovány. Wang et al. (2011) totiž nedokazují homeotický posun jako takový, jen to, že ptačí prsty mají identity I, II, III, což se pod axis-shiftem Towerse a spol. rozumí tak nějak samo sebou – jaké by taky měly mít jiné, když vyrůstají z pozic 1, 2, 3. Přesto si jejich studie zaslouží určitou minimální pozornost. Autoři v ní nasazují postup zvaný transkriptomika, který si na rozdíl od genomiky (zabývající se DNA) a proteomiky (zabývající se veškerými molekulami proteinů, které lze nalézt uvnitř jedné buňky) všímá molekul mRNA, do kterých je genetická informace obsažená v DNA transkribována. Zabývá se tedy především tím, které součástí genomu jsou aktivovány a kdy se tak děje. Autoři tedy vzali prsty z předních a zadních končetin kuřat ve dvou různých stadiích vývoje (5,5 a 7 dní starých), osekvenovali všechny mRNA transkripty, které v nich našli, rozeznali celkem přes 14 000 genů a nad výslednými daty provedli vícerozměrné škálování (MDS). Vynechali všechny geny, jejichž exprese se mezi přední a zadní končetinou lišila, a zjistili, že zbývající geny se shlukují podle dvou kritérií: jednak podle toho, z jak starého kuřete byly sekvenovány, jednak podle toho, zda se nacházejí v palci nebo zbývajících prstech. Ukázalo se přitom, že palec na noze (prst 1/I) a prst, který je palcem z hlediska teorie homeotického posunu (prst 2/I) sdílejí celou řadu vlastností, včetně toho, že v nich nedochází k expresi genů Hoxd12, Hand2, Zic3 a Lhx9, z nichž ty první dva jsou přitom dobře známými negativními markery nejvíce anteriorního prstu (v lidštině: když nejsou exprimovány, je to palec).
Wang et al. (2011) kupodivu vůbec nebyli schopni identifikovat identity dvou zbývajících prstů na křídle. Povedlo se identifikovat několik genů, které spojují prsty II a III na křídle s prsty III a IV na noze (v tomto pořadí), v obou případech jsou však kontradikovány signálem jiných genů, který křídelní prst II spojuje s pozdním prstem II nohy a křídelní prst III s tímtéž prstem na noze u mladšího exempláře. Autoři si také povšimli, že křídelní prst III je jedinečný tím, že v něm dochází k expresi genu Socs2, což souhlasí s tím, co o něm nahlásili Tamura et al. (2011) – že je celý developmentálně divný a že má zřejmě vlastní, odvozenou identitu. Obecně se prsty na křídle zdají být podstatně více diferenciované než prsty na zadní končetině. Právě tato zjištění (a ne "potvrzení" frame-shiftu, které vyplývá už z toho, že se na studii podíleli Young a Wagner) se zdají být na studii Wangové a spol. nejcennější.
*Witmerův článek v některých místech prostě nejde nemilovat:
There is a sense among some that the embryological signal must be correct because it involves seemingly high-tech bench science and makes reference to hox genes, as opposed to the dust and dirt of paleontology.**Roztomilé je, že autoři podporující homeotický posun a pyramidovou redukci si vyčítají úplně stejnou věc: přehnanou důvěru v to, že několik konkrétních fosilií nám říká, jak přesně transformace pětiprsté ruky na tříprstou probíhala. Galis et al. (2003) – jedni z těch, kteří homeotický posun odmítli a vrátili se k tradičnímu "ti druzí to mají špatně", kde "ti druzí" znamenalo paleontology – přišli s poměrně přesvědčivým argumentem: hlavním důkazem pro to, že teropodi ztratili právě prsty IV a V, je jejich výrazná redukce u herrerasaura a eoraptora. Spousta ceratosaurů (kteří jsou ptákům blíže příbuzní) ale prst IV nijak zvlášť zakrnělý nemá, což by mohlo naznačovat, že Herrerasaurus a Eoraptor jej redukovali nezávisle na ostatních teropodech a jako takoví jsou úplně irelevantní k poznání toho, co se odehrávalo na ptačí linii teropodní evoluce. (Nová hypotéza, podle níž je Eoraptor ve skutečnosti hodně primitivním sauropodomorfem, dává Galisovi a spol. za pravdu. Na druhou stranu se zdá, že redukovat prsty IV a V je spíš jakási obecná tendence všech archosaurů, protože tyto prsty jsou menší i u žijících krokodýlů.) Xu et al. (2009) se zase setkali s výtkou, že jejich model až příliš spoléhá na to, že Limusaurus je v rámci ceratosaurů opravdu tak bazální, jak jej odhalila původní fylogenetická analýza; a že si tedy uchoval dostatek primitivních znaků od společného předka ceratosaurů a tetanur, aby nám mohl říct něco zajímavého o tetanurách samotných (kam patří i ptáci, objekt našeho zájmu). To přitom nemusí být pravda: Andrea Cau jej např. ve své slavné Megamatrici vykryl jako noasaurida, tedy zvíře v rámci ceratosaurů poměrně hluboce usazené a tedy zřejmě i odvozené.
Witmer 2002:20
Zdroje:
- http://dml.cmnh.org/2011Aug/msg00173.html
- http://dml.cmnh.org/2011Aug/msg00175.html
- http://dml.cmnh.org/2011Aug/msg00199.html
- http://dml.cmnh.org/2005Dec/msg00213.html
- http://dml.cmnh.org/2008Nov/msg00134.html
- http://theropoda.blogspot.com/2009/06/limusaurus-inextricabilis-xu-et-al-2009.html
- Burke AC, Feduccia A, Hinchliffe R 1998 Counting fingers of birds and dinosaurs. Science 280(5362): 355a
- Cohn MJ, Lovejoy CO, Wolpert L, Coates MI 2002 Branching, segmentation and the metapterygial axis: pattern versus process in the vertebrate limb. BioEssays 24: 460–5
- Galis F, Kundrát M, Sinervo B 2003 An old controversy solved: bird embryos have five fingers. Trends Ecol Evol 18(1): 7–9
- Gillies CD 1929 The origin of the Os pisiforme. J Anat 63: 380–3
- Hinchliffe JR 1985 “One, two, three” or “two, three, four”: an embryologist’s view of the homologies of the digits and carpus of modern birds. 141–7 in Hecht MK, Ostrom JH, Viohl G, Wellnhofer P, eds. The Beginnings of Birds. Eichstätt: Freunde des Jura-Museums
- Holmgren N 1933 On the origin of the tetrapod limb. Acta Zool Stockholm 14: 185–295
- Holmgren N 1952 An embryological analysis of the mammalian carpus and its bearing upon the question of the origin of the tetrapod limb. Acta Zool Stockholm 33: 1–115
- Kundrát M, Seichert V, Russell AP, Smetana K 2002 Pentadactyl pattern of the avian wing autopodium and pyramid reduction hypothesis. J Exp Zool 294: 152–9
- Langer MC 2004 Basal Saurischians. 25–46 in Weishampel DB, Dodson P, Osmólska H, eds. The Dinosauria 2nd edition. Berkeley (CA): Univ of California Press
- Larsson HC, Heppleston AC, Elsey RM 2010 Pentadactyl ground state of the manus of Alligator mississippiensis and insights into the evolution of digital reduction in Archosauria. J Exp Zool Mol Dev Evol 314B: 571–9
- Larsson HCE, Wagner GP 2002 Pentadactyl ground state of the avian wing. J Exp Zool (Mol Dev Evol) 294: 146–51
- Schestakowa GS 1927 Die Entwicklung des Vogelflügels. Bull Soc Nat Moscou (Biol) 36: 163–210
- Shubin NH, Alberch PA 1986 Morphogenetic approach to the origin and basic organization of the tetrapod limb. Evol Biol 20: 319–87
- Steiner H 1934 Über die embryonale Hand- und Fuß-Skelettanlage bei den Crocodiliern, sowie über ihre Beziehung zur Vogel-Flügelanlage und zur ursprünglichen Tetrapoden-Extremität. Rev Suisse Zool 41: 383–96
- Tamura K, Nomura N, Seki R, Yonei-Tamura S, Yokoyama H 2011 Embryological evidence identifies wing digits in birds as digits 1, 2, and 3. Science 331(6018): 753–7
- Towers M, Signolet J, Sherman A, Sang H, Tickle C 2011 Insights into bird wing evolution and digit specification from polarizing region fate maps. Nature Comms 2: 426
- Vargas AO, Fallon JF 2005 Birds have dinosaur wings: The molecular evidence. J Exp Zool B Mol Dev Evol 304: 86–90
- Vargas AO, Kohlsdorf T, Fallon JF, VandenBrooks J, Wagner GP 2008 The evolution of HoxD-11 expression in the bird wing: insights from Alligator mississippiensis. PLoS ONE 3(10): e3325
- Wagner GP 2005 The developmental evolution of avian digit homology: an update. Theory Biosci 124: 165–83
- Wagner GP, Gauthier JA 1999 1,2,3 = 2,3,4: a solution to the problem of the homology of the digits in the avian hand. Proc Natl Acad Sci USA 96: 5111–6
- Wagner GP, Larsson HCE 2007 Fins and limbs in the study of evolutionary novelties. 49–61 in Hall BK, ed. Fins into limbs: evolution, development, and transformation. Chicago: Univ of Chicago Press
- Wang Z, Young RL, Xue H-L, Wagner GP 2011 Transcriptomic analysis of avian digits reveals conserved and derived digit identities in birds. Nature 477(7366): 583–6
- Watson DMS 1913 On the primitive tetrapod limb. Anat Anzeiger 46: 24–7
- Welten MCM, Verbeek FJ, Meijer AH, Richardson MK 2005 Gene expression and digit homology in the chicken embryo wing. Evol Dev 7(1): 18–28
- Witmer LM 2002 The debate on avian ancestry: phylogeny, function, and fossils. 3–30 in Chiappe LM, Witmer LM, eds. Mesozoic Birds: Above the Heads of the Dinosaurs. Berkeley (CA): Univ of California Press
- Xu X, Clark JM, Mo J, Choiniere J, Forster CA, Erickson GM, Hone DWE, Sullivan C, Eberth DA, Nesbitt S, Zhao Q, Hernandez R, Jia C-K, Han F-L, Guo Y 2009 A Jurassic ceratosaur from China helps clarify avian digit homologies. Nature 459(7249): 940–4
- Young RL, Bever GS, Wang Z, Wagner GP 2011 Identity of the avian wing digits: problems resolved and unsolved. Dev Dynam (Special Issue on Limb Development) 240(5): 1042–53











