Hypotézu, podle které je Triceratops, ikonický rohatý dinosaur z pozdní svrchní křídy Severní Ameriky, pouze subadultním jedincem taxonu Torosaurus, odprezentoval poprvé Scannella (2009) na loňském výročním setkání Společnosti pro paleontologii obratlovců (SVP). Zatímco tehdy však šlo pouze o krátký abstrakt, mezitím už čekal na schválení plnohodnotný článek pro časopis Journal of Vertebrate Paleontology, který se nedávno dočkal publikace. Scannella & Horner (2010) v něm ukazují evidenci pro tvrzení, že v obou případech jde pouze o odlišná ontogenetická stadia jediného taxonu, podpořenou bohatým fotografickým materiálem.
Je užitečné poznamenat, že tato "synonymizace skrze ontogenezi" by představovala jiný druh akce, než např. Mikem Taylorem obhajované přesunutí druhu Brachiosaurus brancai pod "rod" Giraffatitan (Taylor 2009). Něco takového je čistě subjektivní, bez ohledu na to, jak velká fenetická mezera odděluje druhy B. brancai a B. altithorax. Na druhou stranu "zrušení" torosaura ve prospěch triceratopse je založeno na paleobiologických důkazech a není subjektivní, Naopak, je vyvratitelné: kdyby existovaly kompletní ontogenetické série od torosaura i triceratopse a v každé fázi by byly navzájem odlišitelné, nikoho by synonymizovat oba taxony zřejmě nenapadlo.
Autoři nejprve připomínají historii obou taxonů. Jejich popisy od sebe dělí pouhé dva roky (Triceratops s rokem popisu 1889 má prioritu) a v obou případech za nimi stojí známý účastník "války o kosti" Othniel Charles Marsh. Systematicky oba taxony spadají mezi chasmosauriny, tedy ceratopsidy s dlouhým límcem (což ve skutečnosti není příliš přesná charakteristika pro triceratopse). Dělení rohatých dinosaurů do dvou skupin podle délky límce sahá daleko do minulosti (Hatcher et al. 1907), až Sternberg (1949) ale předložil fylogenetickou hypotézu blízkou té dnešní, podle které Triceratops ve skutečnosti spadá hluboko do skupiny s dlouhým límcem. Dodson et al. (2004) odhalili torosaura a triceratopse ve fylogenetické analýze velmi blízko sebe; pouze Nedoceratops byl torosaurovi ještě blíže. Zajímavé je, že Nedoceratops sám byl dlouhou dobu považován za synonymum triceratopse.
Zvláštností triceratopse není jen celkově kratší parietoskvamosální límec, ale i to, že v něm chybějí typická okna. Ta mohou být redukovaná (Arrhinoceratops) nebo naopak hypertrofovaná (Chasmosaurus), v nějakém tvaru a velikosti jsou ale přítomna u všech ostatních ceratopsidů. Torosaurus je naopak stavbou své lebky daleko typičtějším chasmosaurinem: protáhlý límec nese dvě výrazná okna. Rozdíly v lebeční morfologii jsou značné a jednoznačně patrné už na první pohled (viz obrázek pod tímto odstavcem). Zvláštní však je, že ve všem ostatním je anatomie obou taxonů dokonale stejná (Farke 2007). Postkraniální kostra je nerozlišitelná a obě zvířata jsou navíc známa ze stejných vrstev stejných geologických jednotek. Autoři se proto snaží ukázat, že tvar a stavba parietoskvamosálního límce byla během vývoje jedinců silně proměnlivá. Všichni zkoumaní jedinci "torosaurů" vykazovali znaky dospělců, což bylo podpořeno i histologickým rozborem; oproti tomu všichni triceratopsové se zdají být nanejvýš subadultní a jejich ontogenetická trajektorie zřejmě byla zakončena stadiem nesoucím typické znaky "torosaura".


AMNH 5116, lebka mladého triceratopsího dospělce (A) srovnaná s lebkou "torosaura", YPM 1830. Dobře vidět jsou protáhlé skvamosály a dopředu mířící zaočnicové rohy. Spodní čelist lebky AMNH 5116 pochází od jiného exempláře; měřítko v obou případech odpovídá 10 cm. (Modifikováno ze Scannella & Horner 2010: Figure 5)
Autoři uvádějí, že studie je jedním z výsledků desetiletého projektu montanského Museum of the Rockies, který se pokouší zrekonstruovat ekosystém, jaký panoval v místech dnešního souvrstí Hell Creek těsně před koncem křídy. Důraz je v něm kladen především na shromažďování dat z většího množství exemplářů stejného druhu a nové podrobnosti o ontogenezi zdejších taxonů jsou jedním z jeho hlavních přínosů. Triceratops je vůbec nejčastějším zdejším dinosaurem, náleží mu až 40% zde nalézaného dinosauřího materiálu, což zahrnuje mimo jiné třicet lebek. Ty zastupují různá ontogenetická stadia, o čemž svědčí jejich velikost, kolísající od půlmetrových lebek po více než dvoumetrové obry (Horner & Goodwin 2006). Torosaurus je podstatně vzácnější a obecně také větší: jeho lebky se pohybují mezi dvěma a třemi metry délky. Triceratopsův materiál, který shromáždilo Museum of the Rockies, byl roztříděn do čtyř obecných ontogenetických kategorií (mládě, juvenilní, subadultní a dospělý jedinec) podle kritérií, která zformulovali Horner & Goodwin (2006). Ty se neomezují na kompletní lebky; k dovození věku často stačí tvar postorbitálních rohů nebo epokcipitálních kostí (druhotných osifikacím, tvořících jakési "výčnělky" na okraji parietoskvamosálního límce). Zatímco ontogenetické změny v proporcích límce autoři zjišťovali posuvným měřidlem, důležitou roli sehrála i histologická analýza.
Ontogenetické série triceratopse ukazují, že v pozdních fázích vývoje temenní kosti začaly zespoda slábnout v místech, odkud u torosaura známe temenní okna. Hypotéza Scannelly a Hornera, podle které jde o zárodek těchto oken, se zdá být přesvědčivá: rozšiřování parietálních oken v průběhu ontogeneze bylo u ceratopsidů zaznamenáno již dříve (Dodson & Currie 1988; Brown et al. 2009). Přestože Forster (1996) diagnostikovala triceratopse skrze jeho extrémně tlusté a pevné temenní kosti, zdaleka nejde o univerzální stav a v místech, kde mají ostatní chasmosaurini v límci okna, je kost u dospělých triceratopsů často silná pouhých 5 milimetrů. Dokonce i u juvenilních triceratopsů je límec nedaleko bočních okrajů (a asi 5 až 10 cm od zadního okraje) poněkud ztenčený. U větších, subadultních jedinců se tyto ztenčené oblasti rozšířily po spodní straně límce a vytvořily velké jamky, výrazné třeba u exempláře YPM 1823. Tyto jamky se dále rozšířily do velkých, půlkruhovitých prohlubní, přiléhajících až ke středové ose límce, na které srůstají obě temenní kosti. V průběhu růstu se prohlubně vzdálily od bočních okrajů límce. Tloušťka parietálu je značně proměnlivá, největší je blízko zadnímu okraji (2,5 až 3 cm) a pak klesá až na 0,5 cm uvnitř prohlubní. V těchto místech se často kost nedochová, zřejmě kvůli křehkosti dané ztenčením. MOR 1122, nejkompletnější lebka "torosaura" (ano, autoři již v této fázi jeho jméno uvozovkují), vykazuje na levé straně parietální okno s vyvýšeným lemem na zadní straně, který u triceratopsů odděluje prohlubně od zbytku límce. Tloušťka kosti temenní je u téti lebky (v místě vzdáleném 2 cm směrem dopředu od zadního okraje límce) 2,62 cm, což je někde uprostřed mezi typickou hodnotou torosaura - 1 až 2 cm - a triceratopse. Právě díky neobvykle velké tloušťce se zřejmě onen lem zachoval; s postupujícím ztenčováním parietálu u jiných "torosaurů" zanikl. MOR 981, což je jediná další "torosauří" lebka, na které se dochoval celý okraj parietálního okna, žádný podobný zesílený lem na své spodní straně nemá. Šířka okna je přitom u MOR 981 o něco větší. Histologická analýza ztenčujícího se parietálu u mladého dospělce triceratopse ukazuje, že k resorpci (opětovnému vstřebání) kosti a otevírání okna docházelo zespoda i zeshora. Mnohočetné opotřebené povrchy ukazují na rozsáhlou remodelaci během ontogeneze.
Spolu s vývojem parietálních oken se u ceratopsů také prodlužuje kost šupinová, druhá z hlavních složek límce. Nejmenší známá triceratopsova lebka, UCMP 154452, má skvamosály tak krátké, že spíše než jiné chasmosauriny připomíná "krátkolímcé" centrosauriny. U chasmosaurinů se šupinové kosti během vývoje prodlužují, Triceratops je ale pedomorfní a udržuje si juvenilní stav až do dospělosti. Široké a krátké skvamosály byly také důvodem, proč byl Triceratops historicky tak často řazen do centrosaurinů (Hatcher et al. 1907; Lull 1933). U exemplářů, která dokumentují velmi pozdní fázi triceratopsovy ontogeneze - konkrétně jde o MOR 2702 a AMNH 5116 - jsou však patrné protáhlé šupinové kosti, někde uprostřed mezi juvenilním stavem reprezentovaným většinou triceratopsů a morfologií známou od "torosaura". Scannella & Horner (2010) dokonce zahrnuli tabulku, ve které ukazují, jak prodlužování šupinových kostí souvisí s rostoucí délkou parietálu, jeho ztenčováním, změnou textury kolem vznikajících oken a jejich otevíráním. "Torosaurovy" skvamosály pro změnu vykazují trend, ve kterém proporční délka kosti (= poměr délky ku šířce) roste s absolutní délkou. Autoři považují za obzvlášť cenný exemplář "torosaura" s označením SMM P97.6.1 - jeho skvamosál je proporčně širší než u jiných "torosaurů" a také se směrem dozadu zašpičaťuje. V ontogenetické sérii, kterou v tabulce zaranžovali z různých exemplářů triceratopse a "torosaura", jde o prvního představitele "torosauří" fáze vývoje: triceratopsové se pohybují mezi fázemi "baby" (parietál dlouhý 12 cm) až po "young adult" (parietál dlouhý až 83 cm), starší dospělci ("adult") jsou reprezentováni nejprve SMM P97.6.1 a pak i dalšími "torosaury".
Následující kapitola se zabývá nově zrekonstruovaným ontogenetickým spektrem a diskuzí o přechodných znacích. AMNH 5116, lebka zmíněná už v předchozím odstavci, ukazuje na velkého jedince (od špičky rostrálu, což je unikátní kost, která se u ceratopsů přesunula před premaxillu, až po kontakt mezi parietálem a skvamosálem měří rovných 200 cm). Má zúžené skvamosály, poměrně tenký parietál (2,3 cm v místě vzdáleném 2 cm od zadního okraje límce) a kostěná jádra rohů mířící dopředu ukazují na dospělejšího jedince, než vidíme u jiných triceratopsů. Jediný důvod, proč nebyla prohlášena za "torosaura" spočívá jen v nepřítomnosti parietálních oken. Důkladnější prozkoumání ale ukáže ztenčené místo v oblasti "torosaurových" oken, a dokonce i u "torosaurových" lebek je kost po okrajích oken tenčí než jinde. Místo obvyklého zbrázděného povrchu u AMNH 5116 v místě budoucíh oken nastupuje hrubší, oblázková textura. Podobná změna v povrchu kosti byla v místech vyvíjejících se oken zaznamenána i u jiných centrosaurinů (Brown et al. 2009). Zbrázděný povrch límce lze pozorovat u juvenilních triceratopsů, jiných ceratopsů a několika exemplářů torosaura. Souvisí zřejmě s rychlým růstem (Sampson et al. 1997) a to, že se vyskytuje u plně dospělých jedinců (což dokazuje histologie) znamená, že v pozdní fázi vývoje límec najednou začal rapidně růst a měnit se.

Scannella a Horner dokonce nezapomínají ani na nedoceratopse, jehož fylogenetická pozice mezi "torosaurem" a "triceratopsem" v některých kladistických analýzách (Dodson et al. 2004) by mohla být pro sjednocování obou taxonů oříškem - bylo by trochu zvláštní, kdyby byl nezávislý druh svými znaky podobnější jednomu ontogenetickému stadiu jiného druhu než jinému. Jeho materiál, USNM 2412, má za sebou pohnutou taxonomickou historii plnou preokupovaných jmen, během které se postupně stával jediným reprezentantem taxonů Diceratops (Lull 1905; Hatcher et al. 1907), Triceratops hatcheri (Lull 1933; Ostrom & Wellnhofer 1986), Diceratus (Mateus et al. 2008) a konečně Nedoceratops (Ukrainsky 2007). Autoři ale usuzují, že nejde o samostatný druh, dokonce ani v rámci triceratopse: jeho autapomorfické znaky - chybějící nasální roh a malá okna v parietálu - zřejmě vůbec nejsou unikátní. Odchylka od malého nosního rohu některých "torosaurů" není zase tak velká (navíc může jít pouze o tafonomický artefakt; v tom případě by se epinasál - kost tvořící jádro nosního rohu, která se někdy nachází odděleně od nasálů a jindy s nimi pevně srůstá [Horner & Goodwin 2006] - jednoduše nedochoval nebo dizartikuloval) a nevelká okna v parietálů přesně zapadající do Scannellova a Hornerova scénáře jako přechodná fáze mezi typickým triceratopsem a "torosaurem". Další podobný přechod představuje lebka UWGM 732 s velmi tenkým parietálem bez oken, protáhlým skvamosálem a možnou přítomností oken v límci. Chybějící zesílení límce v místě kontaktu mezi oběma laterálně umístěnými skvamosály a mezi nimi sevřeným parietálem ale ukazuje, že spíše než o dospělce "torosaurova" typu jde o mladšího dospělce, reprezentovaného některými triceratopsy.
Dále se práce zabývá epiparietály a episkvamosály: jak už jsem nedávno na blogu vysvětloval, jde o malé kosti po okraji ceratpsidního límce. U juvenilního triceratopse mají trojúhelníkový tvar, poté se zplošťují a včleňují do okraje límce. Scannella & Horner (2010) zpochybňují konstatntní výskyt 5 epiparietálů u triceratopse, který by jej měl odlišovat od "torosaura" (Forster 1996; Farke 2007) a místo toho navrhují rostoucí počet během ontogeneze: MOR 2923 disponuje 6 epiparietály, MOR 981 snad až 10 (Farke 2007) a "torosauří" MOR 1122 až 12 těmito elementy. Stejně tak proměnlivý je počet episkvamosálů: Triceratops bývá obyčejně rekonstruován s čtyřmi nebo pěti, u MOR 1120 to ale mohlo být šest na jednom a sedm na druhém skvamosálu, přičemž 7 je počet, který si udržel i "Torosaurus" (MOR 1122).
Osteohistologický rozbor ukazuje, že zatímco zaočnicové rohy malého juvenilního triceratopse se skládaly výhradně z primární kostní tkáně a byly velmi porézní. U většího juvenilního jedince jsou již v primární tkáni patrné sekundární osteony (pro význam pojmů jako "primární tkáň" a "sekundární osteon" si opět dovolím odkázat na starší článek) a kost je celkově méně řídká. Mladí dospělci, poslední vývojové stadium reprezentované "skutečnými" triceratopsy, mají v rozích remodelovanou, ale stále poněkud houbovitou tkáň bez mnohogeneračních Haversových kanálků. Ty najdeme až u starších dospělců reprezentovaných "torosaury", kteří jsou histologicky "dospělejší" než jedinci dříve považovaní za dospělce triceratopse. Autoři z toho usuzují, že Triceratops není žádnou výjimkou z komplikovaných změn, kterými prochází ceratopsidní lebka během ontogeneze (Horner & Goodwin 2006) a že je oproti ostatním chasmosaurinům pedomorfní: udržuje si juvenilní znaky až do velice pozdního bodu na své ontogenetické trajektorii, kdy konečně dojde k proměně, kterou od jiných chasmosaurinů známe z raného věku. Porozumění triceratopsově ontogenezi je důležité pro jeho paleoekologii a systematiku.
Až dosud tedy studie představila dva typy evidence: morfologickou (ontogenetická série sestavená z desítek exemplářů ukazují prodlužování parietálu, zúžování skvamosálů, zplošťování epokcipitálů a otevírání límcových oken) a histologickou. Autoři ale vědí, co by jejich hypotézu mohlo vyvrátit: např. nalezení jiných přijatelných juvenilních jedinců "torosaura", lišících se od triceratopse. Kandidátem byla např. lebka ANSP 15192, která je poměrně malá (177 cm délky) a popsána jako postrádající epokcipitály. To ale Scannella a Horner nepotvrdili, naopak: lebka jimi disponuje a jejich morfologie je typická. Žádný exemplář "torosaura" nemá zaočnicové rohy zakřivené dozadu, což by bylo dobrým ukazatelem nedospělosti (Horner & Goodwin 2006): v průběhu chasmosaurinní ontogeneze tyto rohy opisují špičkami oblouk a z náklonu dozadu se převrátí dopředu. Ani velikost kupodivu není nijak spolehlivým indikátorem: v connecticutském Yale Peabody Museum jsou hned vedle sebe uloženy dvě lebky triceratopse, z nichž jedna je dlouhá 154 cm a má uzavřené lebeční švy, zatímco ta druhá s délkou 187 cm má švy stále otevřené - spolehlivý doklad nedospělosti.
Pokud je nutné zamítnout existenci mladých jedinců "torosaura", co sexuální dimorfizmus? Navrhli jej Ostrom & Wellnhofer (1990); jde však o jev, který není u dinosaurů nijak zvlášť dobře zdokumentovaný (Padian et al. 2004) a navíc nevysvětluje předvídatelnou povahu ztenčování temenních kostí u triceratopse a vyspělejší histologii zaočnicových rohů "torosaura". Pohalvní dimorfizmus však nelze zamítnout při vysvětlování velikostních rozdílů mezi jedinci ve stejném ontogenetickém stadiu. Zajímavou myšlenkou je, že "Torosaurus" by přece jen mohl být samostatným taxonem, který je pouze od triceratopse neodlišitelný až do pozdní fáze ontogeneze. Opět je však zamítnuta, jednak proto, že oba taxony velkých chasmosaurinů by musely žít na stejných lokalitách ve stejném čase, jednak kvůli "dospělejší" histologii "torosaura".

V závěru autoři předkládají novou diagnózu taxonu Triceratops. Rozeznávají v něm dva validní druhy (na rozdíl třeba od studie Dodson et al. 2004): kromě T. horridus také T. prorsus, a to proto, že jsou od sebe stratigarficky odděleni (Scannella & Fowler 2009). Detaily mají následovat v další práci. Problematický je také druh "Torosaurus" utahensis, který po synonymizaci torosaura s triceratopsem může skončit buď jako nový druh triceratopse nebo arrhinoceratopse. Diagnóza triceratopse zahrnuje protáhlá kostěná jádra zaočnicových rohů, vznikajících raným srůstem prefrontálů s postorbitály; proměnlivý epinasální roh; epinasál přirůstající ke komplexu rostrál-nasál-premaxilla; proměnlivý tvar postfrontální fontanely (opět viz zde) a její občasné úplné uzavření v důsledku srůstu frontálů a parietálů; poměrně krátký a široký parietoskvamosální límec vějířovitého tvaru, v nemž se v dospělosti tvoří okna; zvlnění v místě srůstu obou parietálů, které se už v rané ontogenezi promění v širokou parietální příčku; oválná až téměř kruhová parietální okna; u dospělců límec zesílený podél linií kontaktu mezi parietálem a skvamosálem; zadní a boční okraje límce ornamentovány epiparietály a episkvamosály a epijugály srostlé s komplexem jugál-kvadratojugál.
Synonymizace triceratopse s "torosaurem" snižuje diverzitu ceratopsidů v souvrství Hell Creek. Podobná událost už přitom zastihla tyrannosauridy (Carr 1999) a při hojně medializované synonymizaci druhů Pachycephalosaurus wyomingensis, Dracorex hogwartsia a Stygimoloch spinifer obzvlášť tvrdě také pachycefalosauridy (Horner & Goodwin 2009). Autoři se domnívají, že důvod, proč se extrémně tlustý (až 6 cm) límec triceratopse v pozdní ontogenezi začal velmi rychle natahovat, ztenčovat a vyvíjet parietální okna, spočívá v komunikaci uvnitř druhu a možnosti snadno porovnat stáří různých jedinců. Zahrnutí "torosaura" do triceratopsovy ontogenetické trajektorie umožňuje vysvětlit, proč je z celého souvrství Hell Creek známo pouze několik "torosaurů" - vždy plně dospělých - v porovnání s tak hojným triceratopsem. Zřejmě byla triceratopsova úmrtnost velmi vysoká a jen nemnoho jedinců se dožilo plné dospělosti, nebo dospělci žili odděleně od nedospělých zvířat. Poměrně revoluční důsledky má i poslední věta publikace: snižování počtu svrchnokřídových dinosaurů synonymizacemi skrze ontogenezi znamená, že už před koncem druhohor dinosauří rozmanitost klesala, místo toho, aby rostla. To by učinilo K/P katastrofu o něco méně pozoruhodnou - dosud převládala tendence pokládat maastricht, poslední křídový stupeň, za naprostý vrchol dinosauří diverzity. Přestávali být dinosauři vládci souše ještě před velkým vymíráním?
Zdroje:
Spolu s vývojem parietálních oken se u ceratopsů také prodlužuje kost šupinová, druhá z hlavních složek límce. Nejmenší známá triceratopsova lebka, UCMP 154452, má skvamosály tak krátké, že spíše než jiné chasmosauriny připomíná "krátkolímcé" centrosauriny. U chasmosaurinů se šupinové kosti během vývoje prodlužují, Triceratops je ale pedomorfní a udržuje si juvenilní stav až do dospělosti. Široké a krátké skvamosály byly také důvodem, proč byl Triceratops historicky tak často řazen do centrosaurinů (Hatcher et al. 1907; Lull 1933). U exemplářů, která dokumentují velmi pozdní fázi triceratopsovy ontogeneze - konkrétně jde o MOR 2702 a AMNH 5116 - jsou však patrné protáhlé šupinové kosti, někde uprostřed mezi juvenilním stavem reprezentovaným většinou triceratopsů a morfologií známou od "torosaura". Scannella & Horner (2010) dokonce zahrnuli tabulku, ve které ukazují, jak prodlužování šupinových kostí souvisí s rostoucí délkou parietálu, jeho ztenčováním, změnou textury kolem vznikajících oken a jejich otevíráním. "Torosaurovy" skvamosály pro změnu vykazují trend, ve kterém proporční délka kosti (= poměr délky ku šířce) roste s absolutní délkou. Autoři považují za obzvlášť cenný exemplář "torosaura" s označením SMM P97.6.1 - jeho skvamosál je proporčně širší než u jiných "torosaurů" a také se směrem dozadu zašpičaťuje. V ontogenetické sérii, kterou v tabulce zaranžovali z různých exemplářů triceratopse a "torosaura", jde o prvního představitele "torosauří" fáze vývoje: triceratopsové se pohybují mezi fázemi "baby" (parietál dlouhý 12 cm) až po "young adult" (parietál dlouhý až 83 cm), starší dospělci ("adult") jsou reprezentováni nejprve SMM P97.6.1 a pak i dalšími "torosaury".
Následující kapitola se zabývá nově zrekonstruovaným ontogenetickým spektrem a diskuzí o přechodných znacích. AMNH 5116, lebka zmíněná už v předchozím odstavci, ukazuje na velkého jedince (od špičky rostrálu, což je unikátní kost, která se u ceratopsů přesunula před premaxillu, až po kontakt mezi parietálem a skvamosálem měří rovných 200 cm). Má zúžené skvamosály, poměrně tenký parietál (2,3 cm v místě vzdáleném 2 cm od zadního okraje límce) a kostěná jádra rohů mířící dopředu ukazují na dospělejšího jedince, než vidíme u jiných triceratopsů. Jediný důvod, proč nebyla prohlášena za "torosaura" spočívá jen v nepřítomnosti parietálních oken. Důkladnější prozkoumání ale ukáže ztenčené místo v oblasti "torosaurových" oken, a dokonce i u "torosaurových" lebek je kost po okrajích oken tenčí než jinde. Místo obvyklého zbrázděného povrchu u AMNH 5116 v místě budoucíh oken nastupuje hrubší, oblázková textura. Podobná změna v povrchu kosti byla v místech vyvíjejících se oken zaznamenána i u jiných centrosaurinů (Brown et al. 2009). Zbrázděný povrch límce lze pozorovat u juvenilních triceratopsů, jiných ceratopsů a několika exemplářů torosaura. Souvisí zřejmě s rychlým růstem (Sampson et al. 1997) a to, že se vyskytuje u plně dospělých jedinců (což dokazuje histologie) znamená, že v pozdní fázi vývoje límec najednou začal rapidně růst a měnit se.

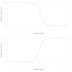
Ontogeneze triceratopsova parietálu. (A) juvenilní jedinec MOR 2951 bez jakéhokoli náznaku budoucího okna, (B) MOR 2924 s nízkou prohlubní na spodním povrchu límce, která nejprve není patrná na jeho horní straně, (C) mladý dospělec MOR 2946 s tenkou oblastí odpovídající budoucímu oknu oddělenou od okolní kosti vyvýšeným lemem, (D) parietální okno staršího dospělce, "torosaura" MOR 1122. (Zdroj: Scannella & Horner 2010: Figure 1)
Scannella a Horner dokonce nezapomínají ani na nedoceratopse, jehož fylogenetická pozice mezi "torosaurem" a "triceratopsem" v některých kladistických analýzách (Dodson et al. 2004) by mohla být pro sjednocování obou taxonů oříškem - bylo by trochu zvláštní, kdyby byl nezávislý druh svými znaky podobnější jednomu ontogenetickému stadiu jiného druhu než jinému. Jeho materiál, USNM 2412, má za sebou pohnutou taxonomickou historii plnou preokupovaných jmen, během které se postupně stával jediným reprezentantem taxonů Diceratops (Lull 1905; Hatcher et al. 1907), Triceratops hatcheri (Lull 1933; Ostrom & Wellnhofer 1986), Diceratus (Mateus et al. 2008) a konečně Nedoceratops (Ukrainsky 2007). Autoři ale usuzují, že nejde o samostatný druh, dokonce ani v rámci triceratopse: jeho autapomorfické znaky - chybějící nasální roh a malá okna v parietálu - zřejmě vůbec nejsou unikátní. Odchylka od malého nosního rohu některých "torosaurů" není zase tak velká (navíc může jít pouze o tafonomický artefakt; v tom případě by se epinasál - kost tvořící jádro nosního rohu, která se někdy nachází odděleně od nasálů a jindy s nimi pevně srůstá [Horner & Goodwin 2006] - jednoduše nedochoval nebo dizartikuloval) a nevelká okna v parietálů přesně zapadající do Scannellova a Hornerova scénáře jako přechodná fáze mezi typickým triceratopsem a "torosaurem". Další podobný přechod představuje lebka UWGM 732 s velmi tenkým parietálem bez oken, protáhlým skvamosálem a možnou přítomností oken v límci. Chybějící zesílení límce v místě kontaktu mezi oběma laterálně umístěnými skvamosály a mezi nimi sevřeným parietálem ale ukazuje, že spíše než o dospělce "torosaurova" typu jde o mladšího dospělce, reprezentovaného některými triceratopsy.
Dále se práce zabývá epiparietály a episkvamosály: jak už jsem nedávno na blogu vysvětloval, jde o malé kosti po okraji ceratpsidního límce. U juvenilního triceratopse mají trojúhelníkový tvar, poté se zplošťují a včleňují do okraje límce. Scannella & Horner (2010) zpochybňují konstatntní výskyt 5 epiparietálů u triceratopse, který by jej měl odlišovat od "torosaura" (Forster 1996; Farke 2007) a místo toho navrhují rostoucí počet během ontogeneze: MOR 2923 disponuje 6 epiparietály, MOR 981 snad až 10 (Farke 2007) a "torosauří" MOR 1122 až 12 těmito elementy. Stejně tak proměnlivý je počet episkvamosálů: Triceratops bývá obyčejně rekonstruován s čtyřmi nebo pěti, u MOR 1120 to ale mohlo být šest na jednom a sedm na druhém skvamosálu, přičemž 7 je počet, který si udržel i "Torosaurus" (MOR 1122).
Osteohistologický rozbor ukazuje, že zatímco zaočnicové rohy malého juvenilního triceratopse se skládaly výhradně z primární kostní tkáně a byly velmi porézní. U většího juvenilního jedince jsou již v primární tkáni patrné sekundární osteony (pro význam pojmů jako "primární tkáň" a "sekundární osteon" si opět dovolím odkázat na starší článek) a kost je celkově méně řídká. Mladí dospělci, poslední vývojové stadium reprezentované "skutečnými" triceratopsy, mají v rozích remodelovanou, ale stále poněkud houbovitou tkáň bez mnohogeneračních Haversových kanálků. Ty najdeme až u starších dospělců reprezentovaných "torosaury", kteří jsou histologicky "dospělejší" než jedinci dříve považovaní za dospělce triceratopse. Autoři z toho usuzují, že Triceratops není žádnou výjimkou z komplikovaných změn, kterými prochází ceratopsidní lebka během ontogeneze (Horner & Goodwin 2006) a že je oproti ostatním chasmosaurinům pedomorfní: udržuje si juvenilní znaky až do velice pozdního bodu na své ontogenetické trajektorii, kdy konečně dojde k proměně, kterou od jiných chasmosaurinů známe z raného věku. Porozumění triceratopsově ontogenezi je důležité pro jeho paleoekologii a systematiku.
Až dosud tedy studie představila dva typy evidence: morfologickou (ontogenetická série sestavená z desítek exemplářů ukazují prodlužování parietálu, zúžování skvamosálů, zplošťování epokcipitálů a otevírání límcových oken) a histologickou. Autoři ale vědí, co by jejich hypotézu mohlo vyvrátit: např. nalezení jiných přijatelných juvenilních jedinců "torosaura", lišících se od triceratopse. Kandidátem byla např. lebka ANSP 15192, která je poměrně malá (177 cm délky) a popsána jako postrádající epokcipitály. To ale Scannella a Horner nepotvrdili, naopak: lebka jimi disponuje a jejich morfologie je typická. Žádný exemplář "torosaura" nemá zaočnicové rohy zakřivené dozadu, což by bylo dobrým ukazatelem nedospělosti (Horner & Goodwin 2006): v průběhu chasmosaurinní ontogeneze tyto rohy opisují špičkami oblouk a z náklonu dozadu se převrátí dopředu. Ani velikost kupodivu není nijak spolehlivým indikátorem: v connecticutském Yale Peabody Museum jsou hned vedle sebe uloženy dvě lebky triceratopse, z nichž jedna je dlouhá 154 cm a má uzavřené lebeční švy, zatímco ta druhá s délkou 187 cm má švy stále otevřené - spolehlivý doklad nedospělosti.
Pokud je nutné zamítnout existenci mladých jedinců "torosaura", co sexuální dimorfizmus? Navrhli jej Ostrom & Wellnhofer (1990); jde však o jev, který není u dinosaurů nijak zvlášť dobře zdokumentovaný (Padian et al. 2004) a navíc nevysvětluje předvídatelnou povahu ztenčování temenních kostí u triceratopse a vyspělejší histologii zaočnicových rohů "torosaura". Pohalvní dimorfizmus však nelze zamítnout při vysvětlování velikostních rozdílů mezi jedinci ve stejném ontogenetickém stadiu. Zajímavou myšlenkou je, že "Torosaurus" by přece jen mohl být samostatným taxonem, který je pouze od triceratopse neodlišitelný až do pozdní fáze ontogeneze. Opět je však zamítnuta, jednak proto, že oba taxony velkých chasmosaurinů by musely žít na stejných lokalitách ve stejném čase, jednak kvůli "dospělejší" histologii "torosaura".

Histologie triceratopsových zaočnicových rohů. (A) juvenilní jedinec UCMP 159233 s houbovitou, vysoce vaskularizovanou kostní tkání a asi 20 cm dlouhým rohem zahnutým dozadu, (B) detail na A ukazující primární tkáň bez sekundárních osteonů, (C) velký juvenilní exemplář MOR-HYP-P31 s dozadu zahnutým, 50 cm dlouhým rohem složeným z houbovité tkáně, (D) detail na C s patrnými řídce rozptýlenými sekundárními osteony, (E) mladý dospělec MOR 1625 s vysoce vaskularizovanou, kompaktní Haversovou kostí, (F) detail na E s dobře viditelnými volně rozmístěnými Haversovými kanálky, (G) mladý dospělec MOR 2702 s podobnou stavbou, jako E, (H) detail na G, (I) "Torosaurus" MOR 1122 s 60 cm dlouhým, dopředu zakřiveným postorbitálním rohem, složeným z husté tkáně s nevelkým prokrvením, (J) detail na I - ve studii zřejmě chybně označeno; viditelné jsou hustě uspořádané Haversovy kanálky. (Zdroj: Scannella & Horner 2010: Figure 7)
V závěru autoři předkládají novou diagnózu taxonu Triceratops. Rozeznávají v něm dva validní druhy (na rozdíl třeba od studie Dodson et al. 2004): kromě T. horridus také T. prorsus, a to proto, že jsou od sebe stratigarficky odděleni (Scannella & Fowler 2009). Detaily mají následovat v další práci. Problematický je také druh "Torosaurus" utahensis, který po synonymizaci torosaura s triceratopsem může skončit buď jako nový druh triceratopse nebo arrhinoceratopse. Diagnóza triceratopse zahrnuje protáhlá kostěná jádra zaočnicových rohů, vznikajících raným srůstem prefrontálů s postorbitály; proměnlivý epinasální roh; epinasál přirůstající ke komplexu rostrál-nasál-premaxilla; proměnlivý tvar postfrontální fontanely (opět viz zde) a její občasné úplné uzavření v důsledku srůstu frontálů a parietálů; poměrně krátký a široký parietoskvamosální límec vějířovitého tvaru, v nemž se v dospělosti tvoří okna; zvlnění v místě srůstu obou parietálů, které se už v rané ontogenezi promění v širokou parietální příčku; oválná až téměř kruhová parietální okna; u dospělců límec zesílený podél linií kontaktu mezi parietálem a skvamosálem; zadní a boční okraje límce ornamentovány epiparietály a episkvamosály a epijugály srostlé s komplexem jugál-kvadratojugál.
Synonymizace triceratopse s "torosaurem" snižuje diverzitu ceratopsidů v souvrství Hell Creek. Podobná událost už přitom zastihla tyrannosauridy (Carr 1999) a při hojně medializované synonymizaci druhů Pachycephalosaurus wyomingensis, Dracorex hogwartsia a Stygimoloch spinifer obzvlášť tvrdě také pachycefalosauridy (Horner & Goodwin 2009). Autoři se domnívají, že důvod, proč se extrémně tlustý (až 6 cm) límec triceratopse v pozdní ontogenezi začal velmi rychle natahovat, ztenčovat a vyvíjet parietální okna, spočívá v komunikaci uvnitř druhu a možnosti snadno porovnat stáří různých jedinců. Zahrnutí "torosaura" do triceratopsovy ontogenetické trajektorie umožňuje vysvětlit, proč je z celého souvrství Hell Creek známo pouze několik "torosaurů" - vždy plně dospělých - v porovnání s tak hojným triceratopsem. Zřejmě byla triceratopsova úmrtnost velmi vysoká a jen nemnoho jedinců se dožilo plné dospělosti, nebo dospělci žili odděleně od nedospělých zvířat. Poměrně revoluční důsledky má i poslední věta publikace: snižování počtu svrchnokřídových dinosaurů synonymizacemi skrze ontogenezi znamená, že už před koncem druhohor dinosauří rozmanitost klesala, místo toho, aby rostla. To by učinilo K/P katastrofu o něco méně pozoruhodnou - dosud převládala tendence pokládat maastricht, poslední křídový stupeň, za naprostý vrchol dinosauří diverzity. Přestávali být dinosauři vládci souše ještě před velkým vymíráním?
Zdroje:
- Brown, C. M., Russell, A. P. & Ryan, M. J. 2009. Pattern and transition of surfacial bone texture of the centrosaurine frill and their ontogenetic and taxonomic implications. Journal of Vertebrate Paleontology 29(1): 132–141.
- Carr, T. D. 1999. Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). Journal of Vertebrate Paleontology 19(3): 497–520.
- Dodson, P. & Currie, P. J. 1988. The smallest ceratopsid skull-Judith River Formation of Alberta. Canadian Journal of Earth Sciences 25: 926–930.
- Dodson, P., Forster, C. A. & Sampson, S. D. 2004. Ceratopsidae. Pp. 494–513 in Weishampel, D. B., Dodson, P. & Osmólska, H. (eds.), The Dinosauria 2nd edition. University of California Press, Berkeley, California.
- Farke, A. A. 2007. Cranial osteology and phylogenetic relationships of the chasmosaurine ceratopsid Torosaurus latus. Pp. 235–257 in Carpenter, K. (ed.), Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis, Indiana.
- Forster, C. A. 1996. Species resolution in Triceratops: cladistic and morphometric approaches. Journal of Vertebrate Paleontology 16(2): 259–270.
- Hatcher, J. B., Marsh, O. C. & Lull, R. S. 1907. The Ceratopsia. US Geological Survey Monograph 49:1–300.
- Horner, J. R. & Goodwin, M. B. 2006. Major cranial changes during Triceratops ontogeny. Proceedings of the Royal Society of London B 273: 2757–2761.
- Horner, J. R. & Goodwin, M. B. 2009. Extreme cranial ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus. PLoS ONE 4(10): e7626.
- Lull, R. 1905. Restoration of the horned dinosaur Diceratops. American Journal of Science 20:420–422.
- Lull, R. S. 1933. A revision of the Ceratopsia or horned dinosaurs. Yale Peabody Museum of Natural History Memoirs 3: 1–178.
- Mateus, O. 2008. Two ornithischian dinosaurs renamed: Microceratops Bohlin, 1953 and Diceratops Lull, 1905. Journal of Paleontology 82: 423.
- Ostrom, J. H. & Wellnhofer, P. 1986. The Munich specimen of Triceratops with a revision of the genus. Zitteliana 14: 111–158.
- Ostrom, J. H. & Wellnhofer, P. 1990. Triceratops: an example of flawed systematics. Pp. 245–254 in Carpenter, K. & Currie, P. J. (eds.), Dinosaur Systematics: Approaches and Perspectives. Cambridge University Press, New York.
- Padian, K., Horner, J. R. & Dhaliwal, J. 2004. Species recognition as the principal cause of bizarre structures in dinosaurs. Journal of Vertebrate Paleontology 24(Supplement to 3): 100A.
- Sampson, S. D., Ryan, M. J. & Tanke, D. H. 1997. Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taxonomic and behavioral implications. Zoological Journal of the Linnean Society 121(2): 293–337.
- Scannella, J. B. 2009. And then there was one: Synonymy consequences of Triceratops cranial ontogeny. Sixty-Ninth Annual Meeting of the Society of Vertberate Paleontology and the Fifty-Seventh Symposium of Vertebrate Palaeontology and Comparative Anatomy (SVPCA), University of Bristol, Bristol, UK, September 23-26, 2009. Journal of Vertebrate Paleontology 29(Supplement to 3): 177A.
- Scannella, J. B. & Fowler, D. W. 2009. Anagenesis in Triceratops: evidence from a newly resolved stratigraphic framework for the Hell Creek Formation. Pp. 148–149 in 9th North American Paleontological Convention Abstracts. Cincinnati Museum Center Scientific Contributions 3.
- Scannella, J. B. & Horner, J. R. 2010. Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny. Journal of Vertebrate Paleontology 30(4): 1157-1168.
- Sternberg, C. M. 1949. The Edmonton fauna and description of a new Triceratops from the Upper Edmonton member; phylogeny of the Ceratopsidae. Annual Report of the National Museum of Canada, Bulletin 113: 33–46.
- Taylor, M. P. 2009. A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914). Journal of Vertebrate Paleontology 29(3): 787-806.
- Ukrainsky, A. S. 2007. A new replacement name for Diceratops Lull, 1905 (Reptilia: Ornithischia: Ceratopsidae). Zoosystematica Rossica 16: 292.