
Když Scannella & Horner (2010) před dvěma lety synonymizovali "torosaura" s triceratopsem na základě argumentu, že morfologické rozdíly mezi těmito dvěma taxony ve skutečnosti pouze odrážejí jiné fáze ontogeneze, vyvolali tím docela pozdvižení.
Jako jeden z prvních jejich závěry explicitně odmítl Longrich (2010) při popisu titanoceratopse, který se odvolával na to, že autoři nedoložili žádný exemplář, jehož morfologie by byla přechodná mezi mladými dospělci (reprezentovanými triceratopsy) a starými dospělci (reprezentovanými "torosaury"). Rozhodl se proto "torosaura" dále rozeznávat coby samostatný taxon, veškeré znaky z jeho revidované diagnózy se ale rekrutovaly právě z té kategorie, kterou Scannella a Horner sáhodlouze diskutovali coby ontogeneticky proměnlivou a tudíž pro podobné účely krajně nespolehlivou. Navíc Longrich neměl zcela pravdu: Scannella a Horner našli exemplář s téměř ideální "tranzitní morfologií", separovaný do samostatného taxonu Nedoceratops. Jeho límec např. vykazuje malá temenní okna, zatímco "Torosaurus" je má daleko širší a límec triceratopse je zcela kompaktní. S tím ale nesouhlasil Farke (2011), podle nějž Nedoceratops skutečně představuje samostatný validní taxon, diagnostikovatelný několika jedinečnými odvozenými znaky.
Celému sporu se dostává dalšího pokračování se dvěma novými studiemi, o nichž píšu v článku níže. Obě vyšly v open-access žurnálu PLoS ONE (hurá!), ta první (Scannella & Horner 2011) dokonce už v prosinci loňského roku. Dnes už bych se o ní asi ani nenamáhal psát, nebýt toho, že na ni na začátku března tak pěkně navázali Longrich & Field (2012) s dosud zřejmě nejúplnější kritikou Scannellovy a Hornerovy hypotézy.
(Drobná poznámka k názvosloví: v článku uvozovkuji jméno "torosaura", zatímco nedoceratopse ponechávám bez uvozovek. Je to poměrně ironické, protože – jak zmiňuji v nadpisu a objasňuji níže – pokud má nějaký z nich skutečně nejisté vyhlídky, je to spíš ten druhý. Jde spíš o věc zvyku: má konvence docela věrně odráží stav, který panoval kolem začátku loňského roku – tehdy poslední slovo ohledně "torosaura" patřilo Scannellovi s Hornerem a ohledně nedoceratopse Farke'ovi. Dnes už tolik smyslu nedává, ale jsem líný ho měnit.)
(Drobná poznámka k názvosloví: v článku uvozovkuji jméno "torosaura", zatímco nedoceratopse ponechávám bez uvozovek. Je to poměrně ironické, protože – jak zmiňuji v nadpisu a objasňuji níže – pokud má nějaký z nich skutečně nejisté vyhlídky, je to spíš ten druhý. Jde spíš o věc zvyku: má konvence docela věrně odráží stav, který panoval kolem začátku loňského roku – tehdy poslední slovo ohledně "torosaura" patřilo Scannellovi s Hornerem a ohledně nedoceratopse Farke'ovi. Dnes už tolik smyslu nedává, ale jsem líný ho měnit.)
1. Scannella & Horner 2011
Je zjevné, že hypotéza Scannelly a Hornera vyžaduje kompletní zavržení taxonu "Torosaurus" - je zbytečné a kontraproduktivní ponechávat si pro jedno a totéž zvíře dvě různá jména a "Torosaurus" musí jít, neboť byl popsán až jako druhý. Populárně naučné články, které o studii referovaly, nicméně trochu pozapomněly na to, že Scannella a Horner neslepili dohromady jen dva, ale hned tři ceratopsidy: tím třetím je výše zmiňovaný Nedoceratops.
Není divu, že úloha tohoto taxonu nebyla příliš zdůrazňována. Nedoceratops je znám z jediné, ne zrovna ideálně zachovalé lebky, a má za sebou pěkně komplikovanou taxonomickou historii. Někteří autoři jej považovali za úplně obyčejného triceratopse (Ostrom & Wellnhofer 1986; Lehman 1998); jiní pro něj v rámci triceratopse vyčlenili vlastní "druh", T. hatcheri (Lull 1933), a ještě další pocítili nutkání dát tomuto "druhu" i zcela nový "rod" (Lull 1905; Forster 1996a). Aby to nebylo tak jednoduché, nedávno se zjistilo, že jméno Diceratops, které pro lebku s katalogovým označením USNM 2412 používala poslední kategorie autorů, už bylo zabráno jakýmsi hmyzem. Bylo tedy nutné přijít se jménem novým, což se skutečně stalo, ale naneštěstí hned dvakrát: Mateus (2008) se pokusil původní název pozměnit na "Diceratus", nebyl si však vědom toho, že o dva měsíce dříve přišel s vlastním návrhem na nové jméno v obskurním ruském žurnálu Ukrainsky (2007). V souladu s principem priority je tak validní Ukrainského název Nedoceratops. (Pokud by čtenáři jméno znělo zvláštně, je pro to dobrý důvod: předpona "nedo-" zde má skutečně stejný význam jako v českých slovech "nedokončený" nebo "nedostatečný" [недостаточный]. Ukrainsky ji zvolil záměrně, neboť se mu lebka zdála na poměry rohatých dinosaurů nedostatečně rohatá.)
Když už tedy víme, jak USNM 2412 říkat v případě, že by si opravdu zasluhovala vlastní taxon, je možné věnovat se otázce, zda si jej skutečně zasluhuje. Její šance vypadají bledě: buď mají Scannella a Horner pravdu, a pak jde prostě o mladého dospělce triceratopse, nebo pravdu nemají – ani pak ovšem Nedoceratops nemusí být validní, neboť navrhované diagnostické znaky mohou být shledány neprůkaznými (viz 2. část tohoto článku) nebo přisouzeny vnitřní variaci v rámci T. horridus (Ostrom & Wellnhofer 1986).
Dosud nejdetailnější obhajobu nedoceratopsovy validity předložil Farke (2011), který zároveň napadl platnost Scannellovy a Hornerovy hypotézy o ontogenetické transformaci triceratopse do "torosaura". Farke dokázal najít celkem tři znaky, které by bylo možné považovat za nedoceratopsovy autapomorfie (tedy jedinečné evoluční novinky). Validita toho zdánlivě nejpřesvědčivějšího z nich, totiž přítomnosti malých oken v kostěném límci, je ovšem velice problematická – ať už Scannella a Horner pravdu mají (viz bod 1.1), nebo nemají (bod 2.3). Na chudáka nedoceratopse tedy útočí obě strany sporu o "torosaurovu" validitu: útok ze strany jejích odpůrců vedou Scannella a Horner (2011), snad i proto, že jsou na této straně debaty zatím jediní.
1.1 Okna v kostěném límci
Přestože rozdíl mezi kompaktním límcem triceratopse a proděravělým límcem "torosaura" a nedoceratopse patří k těm nejnápadnějším, Scannellova a Hornerova hypotéza se s ním paradoxně dokáže vyrovnat nejsnáze. Už v samých počátcích diskuze totiž oba autoři poukázali na skutečnost, že některé exempláře triceratopse mají v límci prohlubně přesně v těch místech, kde u "torosaura" nacházíme okna. Podle nich takto začínal proces resorpce kosti, který nakonec límec zcela perforuje. Je-li to tak, měli bychom dříve či později nějaký exemplář přistihnout uprostřed procesu perforace: od centrosaurinů víme, že temenní okna začínají coby malé otvůrky a s dalším stárnutím zvířete se postupně rozšiřují (Sampson et al. 1997; Brown et al. 2009). Zdá se, že Nedoceratops skutečně přistižen byl, jelikož okno už sice má, ale mimořádně malé, zabírající méně než 5% plochy temenní kosti. (Skutečně jen jedno "okno", jelikož USNM 2412 téměř zcela postrádá levou stranu límce.)
Farke (2011) tuto interpretaci zavrhl, protože kolem okna nedokázal rozpoznat žádnou prohlubeň - ačkoli právě v ní by se podle výše uvedeného scénáře měl otvor začít formovat. Scannella & Horner (2011) nicméně upozorňují, že temenní kost USNM 2412 je v daném místě nejen extrémně tenká, ale na spodní straně také lehce prohloubená. Prohlubeň je sice špatně viditelná a částečně skrytá za kovovou podpěrou, ale při detailním ohledání je její přítomnost patrná (viz fotografii pod tímto odstavcem). V daném místě má navíc kost trochu jinou texturu povrchu.

1.2 Nosní roh
Farke (2011) poukázal na to, že u USNM 2412 lze jen špatně určit místo, kde nosní kost přechází v jádro nasálního rohu, zatímco u ostatních ceratopsidů je toto jádro zcela oddělené. Scannella & Horner (2011) ale podotýkají, že podobnou morfologii nosního rohu vidíme i na lebkách mnohých triceratopsů – vyjmenovávají 5 různých exemplářů a zdůrazňují nevýrazný kostěný hrbolek na MOR 981, který má ke stavu pozorovanému u nedoceratopse obzvlášť blízko (ačkoli jej zcela nedosahuje). To není příliš překvapivé: už dlouho se ví, že tvar a rozměry kostěné základny pro nosní roh jsou u triceratopse značně proměnlivé. Taxon "Ugrosaurus", jehož diagnóza z větší části stála právě na tomto rysu (Cobabe & Fastovsky 1987), byl nedlouho po svém popisu zavržen coby mladší synonymum triceratopse (Forster 1993). Je rovněž možné, že Nedoceratops oddělené kostěné jádro nosního rohu (neboli epinasál) měl a ztratil jej v průběhu života nebo fosilizace (Ostrom & Wellnhofer 1986; Forster 1996a). Farke sice ztrátu rohu za života zvířete zamítl kvůli chybějícímu otevřenému švu, který by v takovém případě šlo očekávat, avšak Scannella a Horner připomínají, že epinasály přirůstají k nosním kostem až poměrně pozdě. Je možné, že nosní kosti samotné srostly až po ztrátě epinasálu, a pak by nebyl důvod čekat otevřený šev.
1.3 Zakřivení zaočniových rohů
Už bylo řečeno, že levá strana lebky není moc dobře zachována; dochovala se na ní ovšem základna zaočnicového rohu, která se zdá být lehce nahnutá směrem dopředu. Pravý roh se zdá být kompletnější a je orientován trochu jinak, více vzpřímeně. Scannella a Horner se však ptají, na kolik je sklon zaočnicových rohů spolehlivým diagnostickým znakem – v průběhu triceratopsovy ontogeneze se totiž radikálně měnil (Horner & Goodwin 2006), což platí i tehdy, když do ní nebudeme začleňovat kontroverzní stadium "torosaura". Mláďata začínala se vzpřímenými růžky, které se později začaly ohýbat dozadu. Scannella a Horner přiznávají, že stupeň zakřivení pozorovatelný u USNM 2412 naznačuje, že lebka patřila poměrně starému jedinci, což není zcela v souladu s jejich hypotézou, podle kterých by měla reprezentovat přechodnou fázi mezi mladými triceratopsy a starými "torosaury". Autoři se však odvolávají na určitý stupeň individuální variace, kterou by snad šlo v případě takto dramatických ontogenetických změn očekávat. Nejde o pouhou výmluvu; Lehman (1990) zdokumentoval, že chasmosaurini (což je podskupina odvozených rohatých dinosaurů charakteristická dlouhými límci, do níž patří i Triceratops) v orientaci kostěné báze postorbitálních rohů skutečně určité "vnitrodruhové" odchylky vykazují.
1.4 Ostatní odlišnosti oproti triceratopsovi
Body 1.1 až 1.3 sice adresují všechny jedinečné znaky, kterými Farke (2011) podpořil nedoceratopsovu validitu, nevyčerpávají však celý seznam znaků, který USNM 2412 odlišuje od všech známých exemplářů triceratopse a "torosaura". Ačkoli tyto znaky již nejsou unikátní (a ani nutně odvozené), představují další překážku pro synonymizaci těchto tří taxonů. Farke (2011) zmiňuje, že dolní okraj kosti šupinové (jedné z dvou kostí, které tvoří typický límec rohatých dinosaurů) se u nedoceratopse nachází vysoko nad alveolárním výběžkem horní čelisti (maxilly), zatímco u triceratopse je tomu naopak. Od "torosaura" nedoceratopse odlišují nejen daleko menší temenní okna (viz výše), ale i nižší počet episkvamosálů, drobných kostí "přilepených" k boční straně kostí šupinových a dodávajících límci zubatý okraj. Farke (2011) rovněž konstatoval, že USNM 2412 má kosti šupinové bez zesíleného mediálního okraje a s redukovaným "jugálním zářezem", což je jakýsi výkus, který najdeme na skvamosálech rohatých dinosaurů naproti kosti čtvercové (Dodson et al. 2004).
Scannella & Horner (2011) ovšem oponují i těmto tvrzením. Na fotografii, kterou jsem přetiskl pod tímto odstavcem, je zřetelně vidět, že exemplář triceratopse s označením USNM 1201 má dokonce vertikální mezeru mezi šupinovou kostí a výběžkem maxilly ještě vyšší, a není sám: UCMP 113697 také vykazuje stav, který Farke popsal od nedoceratopse. Také velikost jugálního zářezu je problematická. U různých exemplářů triceratopse vykazuje nezanedbatelné odchylky a na jedné straně nedoceratopsovy lebky je větší než na té druhé, třebaže patrně v důsledku patologie. Autoři také předpokládají, že zesílení šupinových kostí podél jejich vzájemného kontaktu se objevuje až u starých dospělců triceratopse, tedy "torosaurů", a pak není překvapivé, že tento znak nepozorujeme u nedoceratopse, který by podle jejich hypotézy měl být dospělcem mladým.

Poslední zbývající překážkou pro prohlášení USNM 2412 za triceratopse jsou počty episkvamosálů. V článku o Farkeho studii jsem už zmínil, že paleontologové se specializací na rohaté dinosaury se už dlouho přou o tom, zda je množství těchto dermálních kostí v průběhu života proměnlivé. Tato hypotéza má své zastánce (Forster et al. 1993; Godfrey & Holmes 1995) i odpůrce (Farke 2011; viz rovněž níže). Scannella a Horner pochopitelně musí stát na straně zastánců: pokud z triceratopse skutečně vyrostl "Torosaurus", muselo zvíře v průběhu života 4 až 7 těchto kostí). I jejich hypotéza s sebou ale nese konkrétní předpovědi: pokud Nedoceratops skutečně stál v ontogenetické trajektorii triceratopse tam, kam ho oba autoři umisťují, musel těchto kostí vykazovat nějaký konkrétní počet, a Farke (2011) tvrdil, že odhadovaný počet 5 episkvamosálů je příliš nízký. "Torosauří stadium", které mělo následovat, má totiž těchto elementů sedm. Jeho závěry ale provází nejistota. Farke (2011) přiznal, že přesně určit počet nedoceratopsových episkvamosálů není snadné, jelikož ty přední přirostly k příslušným šupinovým kostem tak dokonale, že je téměř nelze rozeznat. Scannella a Horner se domnívají, že mají lepší interpretaci: nejde je rozeznat, protože tam žádné nejsou. Episkvamosály jsou často ztraceny v průběhu fosilizace (Horner & Goodwin 2008) a USNM 2412 nemusela být v tomto ohledu žádnou výjimkou.
Vzhledem k těmto faktům autoři podotýkají, že dokonce i kdyby Triceratops a "Torosaurus" synonymní nebyli, jediné, co by podporovalo nedoceratopse jako samostatný taxon, jsou jeho malá temenní okna. Jejich přítomnost je ale snadno vysvětlitelná a dokonce očekávatelná, pokud synonymitu obou taxonů připustíme. V další sekci své studie Scannella a Horner rozvíjejí argumenty, proč bychom to měli udělat.
1.5 Triceratops = "Torosaurus"
Ačkoli šlo ve Farkeho studii spíše o vedlejší téma, autor se zapojil i do kontroverze okolo validity "torosaura". Poukázal na odlišné počty episkvamosálů a epiparietálů, na fakt, že temenní okna se v ontogenezi rohatých dinosaurů otevírala poměrně brzy, a ne pozdě, když už zvíře prakticky přestalo růst (největší exempláře triceratopse bez oken jsou stejně velké jako "torosauři" s okny), a na skutečnost, že textura povrchu nedoceratopsova límce ani náhodou neodpovídá tomu, co bychom od "přechodného stadia" mezi oběma zvířaty očekávali.
1.5.1 Epiosifikace
Scannella & Horner (2011) nejprve komentují diskrepance v počtu epiosifikací (což je souhrnný název pro epiparietály a episkvamosály). Dávají Farkemu za pravdu, že i čerstvě vylíhlí triceratopsové mají po okrajích límce 5 až 7 vroubků, což je stejný počet, který nacházíme i u největších exemplářů. Současně však připomínají, že stejné číslo vykazují i MOR 1122 a MPM VP6841, což jsou ovšem "torosauři". Scannella & Horner (2010) nicméně objevili exempláře, kteří mají na levé straně límce jiný počet epiosifikací než na pravé, a už bylo zmíněno, že jejich ztráty kvůli tafonomickým příčinám také nejsou výjimkou. Farke (2011) také podotkl, že u centrosaurinů žádné další epiosifikace v průběhu ontogeneze nepřibývají, což je tvrzení, které autoři napadají hned dvěma způsoby. Jednak je pochybné, zda je srovnání relevantní a límec chasmosaurinů (jako byl Triceratops) se vyvíjel přesně stejně jako u centrosaurinů, jednak i Farke sám ukázal, že počty těchto elementů se na centrosaurinních límcích liší – pouze to přisoudil individuální variaci místo variace ontogenetické.
Oproti své minulé studii (Scannella & Horner 2010) ale autoři trochu modifikují mechanizmus, kterým k přibývání epiosifikací mělo docházet. Místo formování nových a nových elementů v průběhu ontogeneze nyní navrhují, že se každá kost postupně protahovala a nakonec rozdělila na dvě poloviny. To, že morfologie těchto kostí se docela dramaticky měnila, je zdokumentováno dobře: Forster (1996b) dokonce nalezla exemplář, kterému všechny epiosifikace splynuly do jednoho souvislého lemu okolo límce. Pro hypotézu Scannelly a Hornera je důležitější spíše jedinec s označením MOR 2975, jehož episkvamosál je výrazně protáhlý a vykazuje dvě oddělené špičky; podle autorů tak byl přistižen právě uprostřed procesu dělení. Pokud by se tímto způsobem rozdělilo všech 6 epiparietálů, které běžní triceratopsové vykazují, výsledkem by byl přesně stejný počet skvamosálů, jaký nacházíme u "torosaura". Osobně jsem k tomuto argumentu poněkud skeptický: byl-li takový proces opravdu běžnou součástí triceratopsova životní cyklu, měli bychom od něj mít víc než jeden příklad. Ačkoli se takový proces dokáže poněkud lépe vypořádat s faktem, že nikde nevidíme triceratopse, kteří by byli co do počtu epiosifikací někde uprostřed mezi běžnými jedinci a "torosaury", což Scannellovi a Hornerovi vytkli Longrich a Field (viz níže), evidence pro něj se zdá být spíše slabá.
Sami autoři ale upozorňují, že nejde o jediné vysvětlení variace v počtu epiosifikací. Významnou roli mohou hrát i časové mezery mezi různými exempláři. MOR 1122, "Torosaurus" s 12 epiparietály, pochází z velmi nízko uložených vrstev souvrství Hell Creek, kde se žádný jiný exemplář triceratopse (myšleno včetně případných "torosaurů") nevyskytuje. Další "Torosaurus", MOR 981, se nachází ve svrchnějších vrstvách a epiparietálů vykazuje už jen 10. MPM VP6841, který se z této trojice nachází v souvrství vůbec nejvýš, sice nemá sadu epiparietálů kompletní, ale délka nejkompletnějšího elementu nasvědčuje tomu, že jich límec nemohl nést víc než 6 – což je číslo, které se shoduje s klasickými triceratopsy. Autoři se chtějí vyrovnat i s dosud nepříliš zdůrazňovaným aspektem celého problému s límcovými osifikacemi: Triceratops jednu má přímo na styku temenní a šupinové kosti, o "torosaurovi" se to ale nevědělo. Scannella a Horner nicméně zmiňují hned dva jedince, které touto kostí prokazatelně disponovali: u jednoho ji přímo najdeme, u druhého se nedochovala, ale její přítomnost je evidentní ze stop po vyživujících cévách.
1.5.2 Textura
Dalším argumentem pro ontogenetický přechod od triceratopsí morfologie k té "torosauří" je textura povrchu límce. Od centrosaurinů (kteří jsou, jak už bylo zmíněno, poněkud nejistou analogií chasmosaurinů) víme, že v průběhu života se postupně vystřídají tři odlišné textury: "dlouhozrnná" neboli rýhovaná u juvenilních jedinců, "strakatá" u jedinců subadultních a drsná nebo naopak hladká u dospělců. Předpokládá se, že tyto odlišné textury docela spolehlivě korelují s dosaženým ontogenetickým stadiem, neboť vznikají při procesech kostní remodelace, k níž dochází po celou dobu života zvířete. Scannella a Horner poznamenávají, že exempláře "torosaura" se na základě tohoto kritéria zdají vykazovat slušné věkové rozmezí: MOR 981 má na většině límce dlouhozrnnou texturu, temenní kost MOR 1122 je drsná. Histologická analýza však ukazuje, že i límec MOR 981 je tvořen extrémně hustou kostí s mnoha Haversovými kanálky, která indikuje stupeň dospělosti srovnatelný s MOR 1122. Autoři texturu límce coby indikátor relativní dospělosti zcela nezavrhují, navrhují však, že k rychlé expanzi temenních kostí (spojených s dlouhozrnnou texturou) docházeli i u poměrně starých, plně dospělých jedinců, což je konzistentní s hypotézou, že "torosauří" morfologie se začala formovat teprve u dospělců triceratopse.
Scannella & Horner (2010) si povšimli, že u AMNH 5116, zřejmě jednoho z nejstarších exemplářů nesoucích triceratopsí morfologii, nese jinou texturu ztenčující se kost v oblasti údajného budoucího temenního okna a jinou zbytek parietálu. Farke (2011) podotkl, že proces resorpce kosti vůbec nemusí souviset s postupující perforací límce, a Scannella s Hornerem mu dávají za pravdu: jde prostě o doprovodný jev k růstu a remodelaci kostní tkáně. Obracejí však tento fakt proti Farkemu, když píšou, že drsný povrch límce a zcela chybějící dlouhozrnná textura nemohou být použity jako důkaz o plné dospělosti nedoceratopse. Pokud texturu vytváří resorpce, je možné, aby se z té "dospělé", tj. hrubé, druhotně opět stala "juvenilní" dlouhozrnná. Pak ovšem nemůžeme mít jistotu, že by se s límcem nedoceratopse už nic zajímavého nedělo, kdyby zvíře žilo dál.

1.5.3 Temenní okna
Postupný vznik temenních oken je zajímavější než textura okolní kosti. Jsou skutečně ztenčená místa na límci některých triceratopsů předchůdcem výrazných otvorů u "torosaura"? Podle Scannelly a Hornera se nacházejí na stejných místech (což ovšem zpochybňují Longrich & Field [2012], viz níže) a šlo by tedy očekávat kladnou odpověď. Farke (2011) však navrhl, že jde o pouhá místa úponu krčních svalů, která se také nacházejí na podobném místě (Tsuihiji 2010). Autoři uznávají, že tuto hypotézu je obtížné vyvrátit. Mohli bychom se ptát na přítomnost zvláštních rysů kostní mikrostruktury, které souvisí s nepřímým svalovým úponem; svaly se ale stejně dobře mohly připojovat rovnou na okostici, což je hypotéza, která se histologickými daty ověřuje o dost hůře. Podle Scannelly a Hornera ale takový typ úponu není příliš pravděpodobný: vývoj, kterým kosti límce procházely, zřejmě v některých fázích vyžadoval kompletní ztrátu periostu, což by pro přímý úpon znamenalo nepřekonatelný problém. Autoři dále konstatují, že ať už se na daná místa krční svaly upínaly nebo ne, ideální "mezičlánek" v perforaci límce stále představují malá temenní okna nedoceratopse. (To však zpochybňují Longrich a Field, jak uvádím níže v článku.)
1.5.4 Mladý "Torosaurus"
Je evidentní, co by hypotézu Scannelly a Hornera zpochybnilo nejlépe: nález nedospělého "torosaura" se všemi charakteristickými znaky přisuzovanými tomuto taxonu, jako je vysoký počet epiosifikací, dlouhý a hranatý límec s protáhlými kostmi šupinovými a plně proděravělá temenní okna, zároveň však s jednoznačnými indikátory nízkého věku – ideálně histologickými. Autoři zdůrazňují, že důkladný, dlouholetý průzkum souvrství Hell Creek, který už přinesl ovoce v podobě vzácných ontogenetických stadií vícero různých dinosauřích taxonů, nedokázal odkrýt žádný nález, který by odpovídal popsané situaci výše. Soudí tedy, že juvenilní "torosauři" buď nebyli k rozeznání od triceratopsů stejného stáří, nebo neexistovali, protože juvenilní "torosauři" triceratopsové byli.
Farke (2011) ovšem tvrdil, že nedospělého "torosaura" známe: mělo by jít o exemplář YPM 1831, který kombinuje rysy typické pro "torosaury" (pochopitelně v čele s rozsáhlými temenními okny) s několika znaky, které zdánlivě vypovídají o poměrně nízkém věku: dlouhozrnná textura, žádné patrné epiosifikace a několik nesrostlých lebečních kostí. Autoři je jeden po druhém vyvracejí: rýhovaná textura se nijak nevylučuje se statusem dospělce, jak ukazuje výše popsaný případ MOR 981; fakt, že epiosifikace nejsou srostlé s příslušnými kostmi límce mnoho neznamená, jelikož ani u mnoha triceratopsových dospělců se nedochovaly všechny, a časová posloupnost uzavírání švů a srůstání kostí se také nezdá být právě spolehlivým indikátorem dospělosti. Scannella a Horner jednak odkazují na svou předloňskou studii, jednak vyjmenovávají exempláře, které buď kombinují velké tělesné rozměry s nesrostlými elementy (např. MOR 2702), nebo naopak malou velikost s dokončenými fúzemi (např. MOR 1120). Pro dospělost YPM 1831 dále svědčí dopředu zahnuté zaočnicové rohy a velmi velké tělesné rozměry.
1.6 Závěr: implikace pro dinosauří systematiku
Už Dodson (1975) ukázal na kachnozobých dinosaurech, že taxony vykazující v průběhu ontogeneze drastické změny morfologie jsou často zavádějící pro odhady dinosauří diverzity. Největší exempláře, patřící největším jedincům, jsou nejvzácnější a mohou být popsány jako samostatné taxony, čistě na základě přítomnosti určitého stavu některých znaků, které jsou v reálu ontogeneticky proměnlivé (Horner et al. 2011). Naopak mláďata si lze splést s trpasličími příbuznými taxonů, které jsou známy jen z koster dospělců: jako klasický případ autoři citují "miniaturní tyrannosauridy" nanotyranna a raptorexe; spory okolo jejich validity se nicméně táhnou dosud (viz bod 2.4), takže spíše než omyly minulosti ilustrují přetrvávající nejistotu. Zásadní každopádně je, že přehnané odhady diverzity v důsledku nerozpoznaných ontogenetických změn vedou k chybným úsudkům o paleoekologii (Dodson 1975).
V případě rohatých dinosaurů by správnost Scannellových a Hornerových zjištění znamenala další snížení počtu taxonů ve vrstvách maastrichtského stáří, tj. nejmladší svrchní křídy. Znamenalo by to, že neptačí dinosauři nevymřeli na vrcholu sil, jak to podávala dinosauří renezance. Skutečný zlatý věk by bylo nutné posunout dál do minulosti, do ranější křídy; směrem ke katastrofě na rozhraní křídy a paleogénu by se pak diverzita už jen snižovala. Ve srovnání s klasickými představami je to scénář poněkud šokující; studie posledních let se ho však zdají stále více potvrzovat (Archibald 1996; Campione & Evans 2011).
Není divu, že úloha tohoto taxonu nebyla příliš zdůrazňována. Nedoceratops je znám z jediné, ne zrovna ideálně zachovalé lebky, a má za sebou pěkně komplikovanou taxonomickou historii. Někteří autoři jej považovali za úplně obyčejného triceratopse (Ostrom & Wellnhofer 1986; Lehman 1998); jiní pro něj v rámci triceratopse vyčlenili vlastní "druh", T. hatcheri (Lull 1933), a ještě další pocítili nutkání dát tomuto "druhu" i zcela nový "rod" (Lull 1905; Forster 1996a). Aby to nebylo tak jednoduché, nedávno se zjistilo, že jméno Diceratops, které pro lebku s katalogovým označením USNM 2412 používala poslední kategorie autorů, už bylo zabráno jakýmsi hmyzem. Bylo tedy nutné přijít se jménem novým, což se skutečně stalo, ale naneštěstí hned dvakrát: Mateus (2008) se pokusil původní název pozměnit na "Diceratus", nebyl si však vědom toho, že o dva měsíce dříve přišel s vlastním návrhem na nové jméno v obskurním ruském žurnálu Ukrainsky (2007). V souladu s principem priority je tak validní Ukrainského název Nedoceratops. (Pokud by čtenáři jméno znělo zvláštně, je pro to dobrý důvod: předpona "nedo-" zde má skutečně stejný význam jako v českých slovech "nedokončený" nebo "nedostatečný" [недостаточный]. Ukrainsky ji zvolil záměrně, neboť se mu lebka zdála na poměry rohatých dinosaurů nedostatečně rohatá.)
Když už tedy víme, jak USNM 2412 říkat v případě, že by si opravdu zasluhovala vlastní taxon, je možné věnovat se otázce, zda si jej skutečně zasluhuje. Její šance vypadají bledě: buď mají Scannella a Horner pravdu, a pak jde prostě o mladého dospělce triceratopse, nebo pravdu nemají – ani pak ovšem Nedoceratops nemusí být validní, neboť navrhované diagnostické znaky mohou být shledány neprůkaznými (viz 2. část tohoto článku) nebo přisouzeny vnitřní variaci v rámci T. horridus (Ostrom & Wellnhofer 1986).
Dosud nejdetailnější obhajobu nedoceratopsovy validity předložil Farke (2011), který zároveň napadl platnost Scannellovy a Hornerovy hypotézy o ontogenetické transformaci triceratopse do "torosaura". Farke dokázal najít celkem tři znaky, které by bylo možné považovat za nedoceratopsovy autapomorfie (tedy jedinečné evoluční novinky). Validita toho zdánlivě nejpřesvědčivějšího z nich, totiž přítomnosti malých oken v kostěném límci, je ovšem velice problematická – ať už Scannella a Horner pravdu mají (viz bod 1.1), nebo nemají (bod 2.3). Na chudáka nedoceratopse tedy útočí obě strany sporu o "torosaurovu" validitu: útok ze strany jejích odpůrců vedou Scannella a Horner (2011), snad i proto, že jsou na této straně debaty zatím jediní.
1.1 Okna v kostěném límci
Přestože rozdíl mezi kompaktním límcem triceratopse a proděravělým límcem "torosaura" a nedoceratopse patří k těm nejnápadnějším, Scannellova a Hornerova hypotéza se s ním paradoxně dokáže vyrovnat nejsnáze. Už v samých počátcích diskuze totiž oba autoři poukázali na skutečnost, že některé exempláře triceratopse mají v límci prohlubně přesně v těch místech, kde u "torosaura" nacházíme okna. Podle nich takto začínal proces resorpce kosti, který nakonec límec zcela perforuje. Je-li to tak, měli bychom dříve či později nějaký exemplář přistihnout uprostřed procesu perforace: od centrosaurinů víme, že temenní okna začínají coby malé otvůrky a s dalším stárnutím zvířete se postupně rozšiřují (Sampson et al. 1997; Brown et al. 2009). Zdá se, že Nedoceratops skutečně přistižen byl, jelikož okno už sice má, ale mimořádně malé, zabírající méně než 5% plochy temenní kosti. (Skutečně jen jedno "okno", jelikož USNM 2412 téměř zcela postrádá levou stranu límce.)
Farke (2011) tuto interpretaci zavrhl, protože kolem okna nedokázal rozpoznat žádnou prohlubeň - ačkoli právě v ní by se podle výše uvedeného scénáře měl otvor začít formovat. Scannella & Horner (2011) nicméně upozorňují, že temenní kost USNM 2412 je v daném místě nejen extrémně tenká, ale na spodní straně také lehce prohloubená. Prohlubeň je sice špatně viditelná a částečně skrytá za kovovou podpěrou, ale při detailním ohledání je její přítomnost patrná (viz fotografii pod tímto odstavcem). V daném místě má navíc kost trochu jinou texturu povrchu.

Spodní strana pravé temenní kosti USNM 2412, holotypu nedoceratopse. Při správném nasvětlení je patrná prohlubeň lemující malé temenní okno (A), konzistentní s hypotézou, že obdobné prohlubně na límci některých starších triceratopsů předcházejí velkým oknům u "torosaurů". (B) Tatáž fotografie s vyznačeným obrysem prohlubně, jejíž rozlohu naneštěstí zčásti zakrývá kovová konstrukce. Kost je v této zóně výrazně tenčí než mimo ní. Pruh měřítka odpovídá 10 cm. (Modifikováno ze Scannella & Horner 2011: Figure 3)
1.2 Nosní roh
Farke (2011) poukázal na to, že u USNM 2412 lze jen špatně určit místo, kde nosní kost přechází v jádro nasálního rohu, zatímco u ostatních ceratopsidů je toto jádro zcela oddělené. Scannella & Horner (2011) ale podotýkají, že podobnou morfologii nosního rohu vidíme i na lebkách mnohých triceratopsů – vyjmenovávají 5 různých exemplářů a zdůrazňují nevýrazný kostěný hrbolek na MOR 981, který má ke stavu pozorovanému u nedoceratopse obzvlášť blízko (ačkoli jej zcela nedosahuje). To není příliš překvapivé: už dlouho se ví, že tvar a rozměry kostěné základny pro nosní roh jsou u triceratopse značně proměnlivé. Taxon "Ugrosaurus", jehož diagnóza z větší části stála právě na tomto rysu (Cobabe & Fastovsky 1987), byl nedlouho po svém popisu zavržen coby mladší synonymum triceratopse (Forster 1993). Je rovněž možné, že Nedoceratops oddělené kostěné jádro nosního rohu (neboli epinasál) měl a ztratil jej v průběhu života nebo fosilizace (Ostrom & Wellnhofer 1986; Forster 1996a). Farke sice ztrátu rohu za života zvířete zamítl kvůli chybějícímu otevřenému švu, který by v takovém případě šlo očekávat, avšak Scannella a Horner připomínají, že epinasály přirůstají k nosním kostem až poměrně pozdě. Je možné, že nosní kosti samotné srostly až po ztrátě epinasálu, a pak by nebyl důvod čekat otevřený šev.
1.3 Zakřivení zaočniových rohů
Už bylo řečeno, že levá strana lebky není moc dobře zachována; dochovala se na ní ovšem základna zaočnicového rohu, která se zdá být lehce nahnutá směrem dopředu. Pravý roh se zdá být kompletnější a je orientován trochu jinak, více vzpřímeně. Scannella a Horner se však ptají, na kolik je sklon zaočnicových rohů spolehlivým diagnostickým znakem – v průběhu triceratopsovy ontogeneze se totiž radikálně měnil (Horner & Goodwin 2006), což platí i tehdy, když do ní nebudeme začleňovat kontroverzní stadium "torosaura". Mláďata začínala se vzpřímenými růžky, které se později začaly ohýbat dozadu. Scannella a Horner přiznávají, že stupeň zakřivení pozorovatelný u USNM 2412 naznačuje, že lebka patřila poměrně starému jedinci, což není zcela v souladu s jejich hypotézou, podle kterých by měla reprezentovat přechodnou fázi mezi mladými triceratopsy a starými "torosaury". Autoři se však odvolávají na určitý stupeň individuální variace, kterou by snad šlo v případě takto dramatických ontogenetických změn očekávat. Nejde o pouhou výmluvu; Lehman (1990) zdokumentoval, že chasmosaurini (což je podskupina odvozených rohatých dinosaurů charakteristická dlouhými límci, do níž patří i Triceratops) v orientaci kostěné báze postorbitálních rohů skutečně určité "vnitrodruhové" odchylky vykazují.
1.4 Ostatní odlišnosti oproti triceratopsovi
Body 1.1 až 1.3 sice adresují všechny jedinečné znaky, kterými Farke (2011) podpořil nedoceratopsovu validitu, nevyčerpávají však celý seznam znaků, který USNM 2412 odlišuje od všech známých exemplářů triceratopse a "torosaura". Ačkoli tyto znaky již nejsou unikátní (a ani nutně odvozené), představují další překážku pro synonymizaci těchto tří taxonů. Farke (2011) zmiňuje, že dolní okraj kosti šupinové (jedné z dvou kostí, které tvoří typický límec rohatých dinosaurů) se u nedoceratopse nachází vysoko nad alveolárním výběžkem horní čelisti (maxilly), zatímco u triceratopse je tomu naopak. Od "torosaura" nedoceratopse odlišují nejen daleko menší temenní okna (viz výše), ale i nižší počet episkvamosálů, drobných kostí "přilepených" k boční straně kostí šupinových a dodávajících límci zubatý okraj. Farke (2011) rovněž konstatoval, že USNM 2412 má kosti šupinové bez zesíleného mediálního okraje a s redukovaným "jugálním zářezem", což je jakýsi výkus, který najdeme na skvamosálech rohatých dinosaurů naproti kosti čtvercové (Dodson et al. 2004).
Scannella & Horner (2011) ovšem oponují i těmto tvrzením. Na fotografii, kterou jsem přetiskl pod tímto odstavcem, je zřetelně vidět, že exemplář triceratopse s označením USNM 1201 má dokonce vertikální mezeru mezi šupinovou kostí a výběžkem maxilly ještě vyšší, a není sám: UCMP 113697 také vykazuje stav, který Farke popsal od nedoceratopse. Také velikost jugálního zářezu je problematická. U různých exemplářů triceratopse vykazuje nezanedbatelné odchylky a na jedné straně nedoceratopsovy lebky je větší než na té druhé, třebaže patrně v důsledku patologie. Autoři také předpokládají, že zesílení šupinových kostí podél jejich vzájemného kontaktu se objevuje až u starých dospělců triceratopse, tedy "torosaurů", a pak není překvapivé, že tento znak nepozorujeme u nedoceratopse, který by podle jejich hypotézy měl být dospělcem mladým.

Nedoceratopsova výjimečnost. (A) levý boční pohled na USNM 1201, nijak kontroverzní triceratopsí lebku, jejíž nejspodnější bod kosti šupinové (vyznačený horní linkou) stojí vysoko nad alveolárním výběžkem horní čelisti (dolní linka). Tento znak měl být přitom unikátní pro nedoceratopse, jehož lebka USNM 2412 (B) je zobrazena z pravého boku a pro účely srovnání převrácena. U triceratopsí lebky je paradoxně ještě výraznější; je dobře patrné, že vertikální mezera mezi oběma elementy je u USNM 1201 ještě větší. Pruh měřítka odpovídá 10 cm. (Zdroj: Scannella & Horner 2011: Figure 5)
Poslední zbývající překážkou pro prohlášení USNM 2412 za triceratopse jsou počty episkvamosálů. V článku o Farkeho studii jsem už zmínil, že paleontologové se specializací na rohaté dinosaury se už dlouho přou o tom, zda je množství těchto dermálních kostí v průběhu života proměnlivé. Tato hypotéza má své zastánce (Forster et al. 1993; Godfrey & Holmes 1995) i odpůrce (Farke 2011; viz rovněž níže). Scannella a Horner pochopitelně musí stát na straně zastánců: pokud z triceratopse skutečně vyrostl "Torosaurus", muselo zvíře v průběhu života 4 až 7 těchto kostí). I jejich hypotéza s sebou ale nese konkrétní předpovědi: pokud Nedoceratops skutečně stál v ontogenetické trajektorii triceratopse tam, kam ho oba autoři umisťují, musel těchto kostí vykazovat nějaký konkrétní počet, a Farke (2011) tvrdil, že odhadovaný počet 5 episkvamosálů je příliš nízký. "Torosauří stadium", které mělo následovat, má totiž těchto elementů sedm. Jeho závěry ale provází nejistota. Farke (2011) přiznal, že přesně určit počet nedoceratopsových episkvamosálů není snadné, jelikož ty přední přirostly k příslušným šupinovým kostem tak dokonale, že je téměř nelze rozeznat. Scannella a Horner se domnívají, že mají lepší interpretaci: nejde je rozeznat, protože tam žádné nejsou. Episkvamosály jsou často ztraceny v průběhu fosilizace (Horner & Goodwin 2008) a USNM 2412 nemusela být v tomto ohledu žádnou výjimkou.
Vzhledem k těmto faktům autoři podotýkají, že dokonce i kdyby Triceratops a "Torosaurus" synonymní nebyli, jediné, co by podporovalo nedoceratopse jako samostatný taxon, jsou jeho malá temenní okna. Jejich přítomnost je ale snadno vysvětlitelná a dokonce očekávatelná, pokud synonymitu obou taxonů připustíme. V další sekci své studie Scannella a Horner rozvíjejí argumenty, proč bychom to měli udělat.
1.5 Triceratops = "Torosaurus"
Ačkoli šlo ve Farkeho studii spíše o vedlejší téma, autor se zapojil i do kontroverze okolo validity "torosaura". Poukázal na odlišné počty episkvamosálů a epiparietálů, na fakt, že temenní okna se v ontogenezi rohatých dinosaurů otevírala poměrně brzy, a ne pozdě, když už zvíře prakticky přestalo růst (největší exempláře triceratopse bez oken jsou stejně velké jako "torosauři" s okny), a na skutečnost, že textura povrchu nedoceratopsova límce ani náhodou neodpovídá tomu, co bychom od "přechodného stadia" mezi oběma zvířaty očekávali.
1.5.1 Epiosifikace
Scannella & Horner (2011) nejprve komentují diskrepance v počtu epiosifikací (což je souhrnný název pro epiparietály a episkvamosály). Dávají Farkemu za pravdu, že i čerstvě vylíhlí triceratopsové mají po okrajích límce 5 až 7 vroubků, což je stejný počet, který nacházíme i u největších exemplářů. Současně však připomínají, že stejné číslo vykazují i MOR 1122 a MPM VP6841, což jsou ovšem "torosauři". Scannella & Horner (2010) nicméně objevili exempláře, kteří mají na levé straně límce jiný počet epiosifikací než na pravé, a už bylo zmíněno, že jejich ztráty kvůli tafonomickým příčinám také nejsou výjimkou. Farke (2011) také podotkl, že u centrosaurinů žádné další epiosifikace v průběhu ontogeneze nepřibývají, což je tvrzení, které autoři napadají hned dvěma způsoby. Jednak je pochybné, zda je srovnání relevantní a límec chasmosaurinů (jako byl Triceratops) se vyvíjel přesně stejně jako u centrosaurinů, jednak i Farke sám ukázal, že počty těchto elementů se na centrosaurinních límcích liší – pouze to přisoudil individuální variaci místo variace ontogenetické.
Oproti své minulé studii (Scannella & Horner 2010) ale autoři trochu modifikují mechanizmus, kterým k přibývání epiosifikací mělo docházet. Místo formování nových a nových elementů v průběhu ontogeneze nyní navrhují, že se každá kost postupně protahovala a nakonec rozdělila na dvě poloviny. To, že morfologie těchto kostí se docela dramaticky měnila, je zdokumentováno dobře: Forster (1996b) dokonce nalezla exemplář, kterému všechny epiosifikace splynuly do jednoho souvislého lemu okolo límce. Pro hypotézu Scannelly a Hornera je důležitější spíše jedinec s označením MOR 2975, jehož episkvamosál je výrazně protáhlý a vykazuje dvě oddělené špičky; podle autorů tak byl přistižen právě uprostřed procesu dělení. Pokud by se tímto způsobem rozdělilo všech 6 epiparietálů, které běžní triceratopsové vykazují, výsledkem by byl přesně stejný počet skvamosálů, jaký nacházíme u "torosaura". Osobně jsem k tomuto argumentu poněkud skeptický: byl-li takový proces opravdu běžnou součástí triceratopsova životní cyklu, měli bychom od něj mít víc než jeden příklad. Ačkoli se takový proces dokáže poněkud lépe vypořádat s faktem, že nikde nevidíme triceratopse, kteří by byli co do počtu epiosifikací někde uprostřed mezi běžnými jedinci a "torosaury", což Scannellovi a Hornerovi vytkli Longrich a Field (viz níže), evidence pro něj se zdá být spíše slabá.
Sami autoři ale upozorňují, že nejde o jediné vysvětlení variace v počtu epiosifikací. Významnou roli mohou hrát i časové mezery mezi různými exempláři. MOR 1122, "Torosaurus" s 12 epiparietály, pochází z velmi nízko uložených vrstev souvrství Hell Creek, kde se žádný jiný exemplář triceratopse (myšleno včetně případných "torosaurů") nevyskytuje. Další "Torosaurus", MOR 981, se nachází ve svrchnějších vrstvách a epiparietálů vykazuje už jen 10. MPM VP6841, který se z této trojice nachází v souvrství vůbec nejvýš, sice nemá sadu epiparietálů kompletní, ale délka nejkompletnějšího elementu nasvědčuje tomu, že jich límec nemohl nést víc než 6 – což je číslo, které se shoduje s klasickými triceratopsy. Autoři se chtějí vyrovnat i s dosud nepříliš zdůrazňovaným aspektem celého problému s límcovými osifikacemi: Triceratops jednu má přímo na styku temenní a šupinové kosti, o "torosaurovi" se to ale nevědělo. Scannella a Horner nicméně zmiňují hned dva jedince, které touto kostí prokazatelně disponovali: u jednoho ji přímo najdeme, u druhého se nedochovala, ale její přítomnost je evidentní ze stop po vyživujících cévách.
1.5.2 Textura
Dalším argumentem pro ontogenetický přechod od triceratopsí morfologie k té "torosauří" je textura povrchu límce. Od centrosaurinů (kteří jsou, jak už bylo zmíněno, poněkud nejistou analogií chasmosaurinů) víme, že v průběhu života se postupně vystřídají tři odlišné textury: "dlouhozrnná" neboli rýhovaná u juvenilních jedinců, "strakatá" u jedinců subadultních a drsná nebo naopak hladká u dospělců. Předpokládá se, že tyto odlišné textury docela spolehlivě korelují s dosaženým ontogenetickým stadiem, neboť vznikají při procesech kostní remodelace, k níž dochází po celou dobu života zvířete. Scannella a Horner poznamenávají, že exempláře "torosaura" se na základě tohoto kritéria zdají vykazovat slušné věkové rozmezí: MOR 981 má na většině límce dlouhozrnnou texturu, temenní kost MOR 1122 je drsná. Histologická analýza však ukazuje, že i límec MOR 981 je tvořen extrémně hustou kostí s mnoha Haversovými kanálky, která indikuje stupeň dospělosti srovnatelný s MOR 1122. Autoři texturu límce coby indikátor relativní dospělosti zcela nezavrhují, navrhují však, že k rychlé expanzi temenních kostí (spojených s dlouhozrnnou texturou) docházeli i u poměrně starých, plně dospělých jedinců, což je konzistentní s hypotézou, že "torosauří" morfologie se začala formovat teprve u dospělců triceratopse.
Scannella & Horner (2010) si povšimli, že u AMNH 5116, zřejmě jednoho z nejstarších exemplářů nesoucích triceratopsí morfologii, nese jinou texturu ztenčující se kost v oblasti údajného budoucího temenního okna a jinou zbytek parietálu. Farke (2011) podotkl, že proces resorpce kosti vůbec nemusí souviset s postupující perforací límce, a Scannella s Hornerem mu dávají za pravdu: jde prostě o doprovodný jev k růstu a remodelaci kostní tkáně. Obracejí však tento fakt proti Farkemu, když píšou, že drsný povrch límce a zcela chybějící dlouhozrnná textura nemohou být použity jako důkaz o plné dospělosti nedoceratopse. Pokud texturu vytváří resorpce, je možné, aby se z té "dospělé", tj. hrubé, druhotně opět stala "juvenilní" dlouhozrnná. Pak ovšem nemůžeme mít jistotu, že by se s límcem nedoceratopse už nic zajímavého nedělo, kdyby zvíře žilo dál.
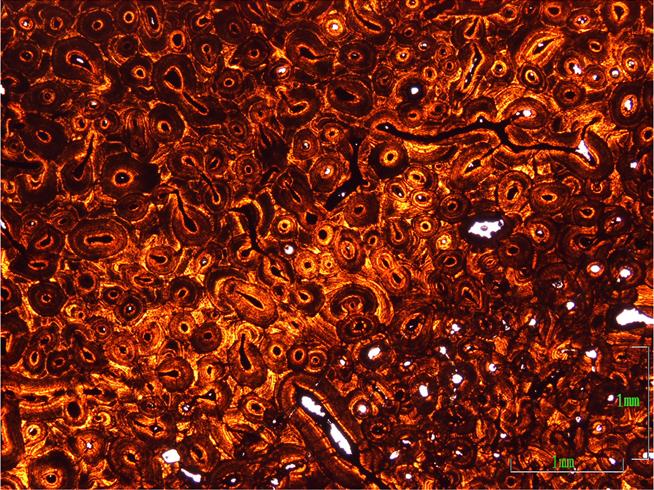
Histologie kostěného jádra zaočnicového rohu MOR 981. Tento exemplář triceratopse vykazuje na kostěném límci stejnou dlouhozrnnou texturu jako údajný subadultní "Torosaurus" YPM 1831, hustá tkáň s mnoha sekundárními osteony (Haversovými systémy) však svědčí o plné dospělosti. Pokud mohou zmiňovanou texturu vykazovat i dospělci, vyvrací to další argument proti postupné přeměně triceratopse v "torosaura". (Zdroj: Scannella & Horner 2011: Figure 8)
1.5.3 Temenní okna
Postupný vznik temenních oken je zajímavější než textura okolní kosti. Jsou skutečně ztenčená místa na límci některých triceratopsů předchůdcem výrazných otvorů u "torosaura"? Podle Scannelly a Hornera se nacházejí na stejných místech (což ovšem zpochybňují Longrich & Field [2012], viz níže) a šlo by tedy očekávat kladnou odpověď. Farke (2011) však navrhl, že jde o pouhá místa úponu krčních svalů, která se také nacházejí na podobném místě (Tsuihiji 2010). Autoři uznávají, že tuto hypotézu je obtížné vyvrátit. Mohli bychom se ptát na přítomnost zvláštních rysů kostní mikrostruktury, které souvisí s nepřímým svalovým úponem; svaly se ale stejně dobře mohly připojovat rovnou na okostici, což je hypotéza, která se histologickými daty ověřuje o dost hůře. Podle Scannelly a Hornera ale takový typ úponu není příliš pravděpodobný: vývoj, kterým kosti límce procházely, zřejmě v některých fázích vyžadoval kompletní ztrátu periostu, což by pro přímý úpon znamenalo nepřekonatelný problém. Autoři dále konstatují, že ať už se na daná místa krční svaly upínaly nebo ne, ideální "mezičlánek" v perforaci límce stále představují malá temenní okna nedoceratopse. (To však zpochybňují Longrich a Field, jak uvádím níže v článku.)
1.5.4 Mladý "Torosaurus"
Je evidentní, co by hypotézu Scannelly a Hornera zpochybnilo nejlépe: nález nedospělého "torosaura" se všemi charakteristickými znaky přisuzovanými tomuto taxonu, jako je vysoký počet epiosifikací, dlouhý a hranatý límec s protáhlými kostmi šupinovými a plně proděravělá temenní okna, zároveň však s jednoznačnými indikátory nízkého věku – ideálně histologickými. Autoři zdůrazňují, že důkladný, dlouholetý průzkum souvrství Hell Creek, který už přinesl ovoce v podobě vzácných ontogenetických stadií vícero různých dinosauřích taxonů, nedokázal odkrýt žádný nález, který by odpovídal popsané situaci výše. Soudí tedy, že juvenilní "torosauři" buď nebyli k rozeznání od triceratopsů stejného stáří, nebo neexistovali, protože juvenilní "torosauři" triceratopsové byli.
Farke (2011) ovšem tvrdil, že nedospělého "torosaura" známe: mělo by jít o exemplář YPM 1831, který kombinuje rysy typické pro "torosaury" (pochopitelně v čele s rozsáhlými temenními okny) s několika znaky, které zdánlivě vypovídají o poměrně nízkém věku: dlouhozrnná textura, žádné patrné epiosifikace a několik nesrostlých lebečních kostí. Autoři je jeden po druhém vyvracejí: rýhovaná textura se nijak nevylučuje se statusem dospělce, jak ukazuje výše popsaný případ MOR 981; fakt, že epiosifikace nejsou srostlé s příslušnými kostmi límce mnoho neznamená, jelikož ani u mnoha triceratopsových dospělců se nedochovaly všechny, a časová posloupnost uzavírání švů a srůstání kostí se také nezdá být právě spolehlivým indikátorem dospělosti. Scannella a Horner jednak odkazují na svou předloňskou studii, jednak vyjmenovávají exempláře, které buď kombinují velké tělesné rozměry s nesrostlými elementy (např. MOR 2702), nebo naopak malou velikost s dokončenými fúzemi (např. MOR 1120). Pro dospělost YPM 1831 dále svědčí dopředu zahnuté zaočnicové rohy a velmi velké tělesné rozměry.
1.6 Závěr: implikace pro dinosauří systematiku
Už Dodson (1975) ukázal na kachnozobých dinosaurech, že taxony vykazující v průběhu ontogeneze drastické změny morfologie jsou často zavádějící pro odhady dinosauří diverzity. Největší exempláře, patřící největším jedincům, jsou nejvzácnější a mohou být popsány jako samostatné taxony, čistě na základě přítomnosti určitého stavu některých znaků, které jsou v reálu ontogeneticky proměnlivé (Horner et al. 2011). Naopak mláďata si lze splést s trpasličími příbuznými taxonů, které jsou známy jen z koster dospělců: jako klasický případ autoři citují "miniaturní tyrannosauridy" nanotyranna a raptorexe; spory okolo jejich validity se nicméně táhnou dosud (viz bod 2.4), takže spíše než omyly minulosti ilustrují přetrvávající nejistotu. Zásadní každopádně je, že přehnané odhady diverzity v důsledku nerozpoznaných ontogenetických změn vedou k chybným úsudkům o paleoekologii (Dodson 1975).
V případě rohatých dinosaurů by správnost Scannellových a Hornerových zjištění znamenala další snížení počtu taxonů ve vrstvách maastrichtského stáří, tj. nejmladší svrchní křídy. Znamenalo by to, že neptačí dinosauři nevymřeli na vrcholu sil, jak to podávala dinosauří renezance. Skutečný zlatý věk by bylo nutné posunout dál do minulosti, do ranější křídy; směrem ke katastrofě na rozhraní křídy a paleogénu by se pak diverzita už jen snižovala. Ve srovnání s klasickými představami je to scénář poněkud šokující; studie posledních let se ho však zdají stále více potvrzovat (Archibald 1996; Campione & Evans 2011).
2. Longrich & Field 2012
V úvodu své studie autoři upozorňují, že pokud by "Torosaurus" a Triceratops skutečně byli různé růstové fáze jediného taxonu, mělo by to dalekosáhlý dopad na dinosauří taxonomii. Bylo by nutné zcela přehodnotit diverzitu rohatých dinosaurů, kde se v současnosti nové taxony rozeznávají právě na základě těch rozdílů, které Scannella a Horner označují za ontogeneticky proměnlivé. Už dnes jsou paleontologové při popisování nových taxonů daleko opatrnější a konzervativnější, než tomu bylo v minulosti (Benton 2008a, b), a synonymizace taxonů zdánlivě tak rozdílných, jako jsou "Torosaurus", Triceratops a Nedoceratops by tento trend jistě ještě posílila. Hypotéza o synonymitě s sebou zároveň nese spoustu testovatelných predikcí, díky čemuž v tomto případě splitting a lumping není otázkou vkusu, ale výzkumu. Longrich a Field formulují následující podmínky, které musí hypotéza o synonymitě splnit: 1) výskyt triceratopse i "torosaura" se musí časově i geograficky překrývat, 2) exempláře obou taxonů musí konzistentně patřit odlišným vývojovým stadiím a 3) musejí existovat nějaké přechodné formy.
2.1 Stratigrafie a biogeografie
Zde hypotéza Scannelly a Hornera na žádné problémy nenaráží. Oba taxony jsou známy ze stejného období (pozdního maastrichtu, tj. nejzazší svrchní křídy) stejné oblasti (západu Severní Ameriky): kostry "torosaura" a triceratopse jsou nalézány ve stejných souvrstvích od Saskatchewanu po Colorado a tam, kde nalézáme jen jednoho z nich (triceratopse v kanadské Albertě, "torosaura" na americkém jihozápadě), klidně může jít o pouhý artefakt nedostatečného fosilního záznamu.

2.2 Relativní věk
Je-li "Torosaurus" starý Triceratops, pak by morfologie obou taxonů měla být omezena na určitý věkový interval: neměli bychom nalézat ani příliš mladé "torosaury", ani příliš staré triceratopse. Scannella & Horner (2010) tuto předpověď otestovali histologickým rozborem zaočnicových rohů, Horner & Lamm (2011) pak tutéž metodu aplikovali na temenní kosti. První studie odhalila, že u "torosaura" došlo k rozsáhlejší kostní remodelaci (tj. odbourávání staré kostní tkáně a jejímu nahrazení novou tkání) než u různých exemplářů triceratopse, což si autoři vyložili jako důkaz o jeho větším stáří. Obě studie naneštěstí od "torosaura" prozkoumaly jediný exemplář, takže jejich závěry jsou statisticky naprosto bezcenné. Je také pochybné, že zrovna míra remodelace kosti by měla spolehlivě indikovat dosažené ontogenetické stadium: Lanyon et al. (1982) a O'Connor & Lanyon (1982) totiž experimentem ukázali, že kost může projít přídavnou remodelací, pokud změníme způsob jejího namáhání. Tempo obměňování kostní tkáně se tedy mění podle toho, jak se mění zátěž, kterou kost prodělává, a liší se nejen kost od kosti, ale dokonce i v rámci kosti jediné podle toho, které místo je namáháno nejvíc.
Longrich & Field (2012) pro svou studii vzorek prozkoumaných "torosaurů" podstatně rozšířili, aby zjistili, zda jsou takto označované formy skutečně konzistentně starší než trcieratopsové. Z výše uvedených důvodů k tomu ovšem nevyužívají Scannellovy a Hornerovy histologické rozbory kostěného límce. Histologie dlouhých kostí, která se pro zjišťování stáří zvířete v době smrti obvykle používá, by byla daleko vhodnější; naneštěstí ale není aplikovatelná, neboť většinu exemplářů "torosaura" známe jen z izolované lebky. Nezbývá tedy, než se pokusit odvodit věk jejich materiálů z údajů nashromážděných od kompletnějších rohatých dinosaurů. Je např. známo, že v průběhu stárnutí těchto zvířat prodělávaly jejich lebky řadu morfologických změn: límec se prodlužoval, zaočnicové rohy se protahovaly a zakřivovaly, povrch kosti se měnil z rýhovaného na drsný a sukovitý. Kosti obklopující očnici srostly s nasály a frontály, rostrální kost (jedinečná vývojová novinka rohatých dinosaurů) přirostla ke kosti předčelistní a ta zase ke kosti nosní. Kosti dermálního původu, tvořící rohy nebo okraje kostěného límce, přirostly k příslušným kostem lebky; srostly i některé elementy mozkovny. To má svůj význam, protože u dnešních savců se takové lebeční srůsty odehrávají v typické sekvenci, z níž lze odvodit stáří zvířete (Wang et al. 2006; Sanchez-Villagra 2010). Konkrétně se zdá, že jakmile jsou jednou fúze kostí završeny a veškeré švy uzavřeny, zvíře dosáhlo plné skeletální dospělosti a růst může už jen velice omezeně. Takový indikátor už byl jednou využit při studiu růstových sérií triceratopse (Fujiwara & Takakuwa 2011) a není tedy důvod, proč by se nemohl uplatnit i u "torosaura".
Autoři proto vybrali 24 ontogeneticky proměnlivých osteologických znaků a podívali se, jaké rozdíly vykazují napříč 36 různými exempláři rohatých dinosaurů. Ty pocházejí od taxonů Triceratops horridus, Triceratops prorsus, "Torosaurus latus", "Torosaurus utahensis" a Tatankaceratops sacrisonorum. Už jen za tímto základním pracovním roztříděním stojí obrovské množství kontroverze: pomineme-li samotné oddělování "torosaura" a triceratopse (prováděné z větší části na základě právě těch kontroverzních znaků, jejichž ontogenetická proměnlivost se teprve má testovat), status "T. utahensis" je velice nejistý a Tatankaceratops je až zarážejícně podobný T. prorsus – až na to, že podivným způsobem míchá znaky typické pro mladá a dospělá zvířata. Podle Longriche (2010) jde buď o exemplář prorsa, který přestal růst ještě před dosažením běžné velikosti, nebo nový, trpasličí subtaxon triceratopse. Zvláštní zmínku si zaslouží Nedoceratops, který je prý ve skutečnosti jen poškozeným exemplářem T. horridus: díry v límci jsou patologické a nejde o pravá temenní okna (vzhledem k jejich nepravidelnému tvaru), zaočnicový roh kolmý na zubní řadu pak lze údajně nalézt jen na jedné straně lebky a jde tak jen o tafonomický artefakt. Zdá se tedy, že už tu máme minimálně tři konfliktní interpretace USNM 2412: Longrich s Fieldem a Scannella s Hornerem se shodnou na tom, že jde o triceratopse (contra Farke 2011), ne už ale na tom, zda je jeho divná lebka jedinečnou ukázkou "přechodné morfologie", nebo jen trochu rozmáčknutá. Ať tak či onak, nashromážděná ontogeneticky informativní data autoři podrobili rozboru metodou maximální parsimonie, která vzhledem k velkému množství chybějících dat a malé variabilitě našla extrémně velké množství optimálních stromů. Autoři ji nechali běžet jen do 250 000 stromů, z nichž byl následně vytvořen striktní konsenzus přetištěný níže (tj. strom obsahující jen ty shluky, které byly přítomné na každém stromě).

Výsledný strom v tomto případě nereprezentuje průběh fylogeneze, nýbrž ontogeneze: jedinci posazení na jeho bázi jsou nejmladší, ti na "vršku" zase nejstarší. Model to samozřejmě není ideální: ačkoli je datová matrice malá, vyskytly se v ní znakové konflikty a problémem jsou i již zmiňovaná chybějící data. Přesto jsou na výsledném stromě rozeznatelné tři odlišné fáze, které Longriech a Field ztotožňují s mláďaty, subadultními jedinci a dospělci. V první fázi se protáhly a rozšířily temenní kosti a zaočnicové rohy se prodloužily, zmohutněly a zakřivily. V druhé fázi spolu navzájem srostly kosti nosní, slzné, předčelní, čelní a zaočnicové a několik kostí mozkovny; textura tvářových kostí a límce začala pomalu hrubnout. Překvapivé je, že v této fázi najdeme největší zdokumentovaná zvířata vůbec. Patří sem totiž i již zmiňovaný "subadultní torosaur" YPM 1831, vůbec největší zástupce svého taxonu, nebo lebka triceratopse s katalogovým označením YPM 1821, která s délkou 101 cm o plných 15 cm přesahuje lebky některých dospělců. (Longrich a Field ujišťují, že podobné nesrovnalosti mezi tělesnými rozměry a dosaženým stupněm ontogeneze panují i u triceratopse.) Konečně během třetí fáze přirostly epokcipitály k límci, epijugály ke kosti jařmové a nosní kost srostla s premaxillou a rostrálem. Autoři také zaznamenali jisté individuální odchylky: změna textury zřejmě u některých jedinců nastala později než u jiných. Matoucí znakové konflikty nastávají i jinde: YPM 1831 nemá srostlý týlní hrbol – na rozdíl od zvířat, která se podle ostatních znaků zdají být mladší – a Tatankaceratops směšuje znaky juvenilních a nedospělých jedinců.
V souvislosti s vyvracením Scannellovy a Hornerovy hypotézy je ale nejzajímavější to, kde se umístilo šest prozkoumaných "torosaurů". Tři z nich se zařadili do kategorie dospělců, minimálně jeden z nich se však jeví mladší, než mnohé exempláře triceratopse. Dva další (včetně tolik problematického YPM 1831) stojí na rozmezí kategorie dospělců a subadultních jedinců; YPM 1831 se nakonec skutečně zdá být spíše subadultní, jak o něm napsal Farke (2011). Důležité ale je, že analýza Longriche a Fielda tento závěr podložila nezávislou evidencí a subadultní status pro YPM 1831 není založen na znacích, které kritizovali Scannella & Horner (2011; viz výše). Místo toho jej podporuje volný epijugál, rostrální kost nepřirostlá k premaxille a již zmiňovaný nesrostlý týlní hrbol; tedy znaky, které u dospělců rohatých dinosaurů nejsou obvyklé. Je pravda, že ani sekvence fúzí lebečních kostí není podle Scannelly a Hornera ontogeneticky informativní; s touto námitkou se však autoři už vypořádali výše. Subadultním se dále zdá být i "Torosaurus" utahensis. Analýza naopak odhalila poměrně velké množství (10) dospělých triceratopsů, což ostře kontrastuje s výsledky Scannelly a Hornera (2010), kteří nebyli schopni nalézt ani jednoho. Podle Longriche a Fielda je to způsobeno tím, že předchozí dvojice autorů zařadila do své studie jen dva dostatečně velké triceratopsí exempláře. Žádný juvenilní "Torosaurus", který by Scannellovu a Hornerovu hypotézu vyvrátil nejlépe, v analýze identifikován nebyl.

2.3 Přechodné formy
Pokud mají Scannella a Horner pravdu a triceratopsi svůj život končili s "torosauří" morfologií lebky, musela jejich lebka v průběhu života absolvovat výrazné morfologické změny: límec se měl protáhnout a změnit tvar, resorpcí kostní tkáně měla vzniknout temenní okna a na jeho okrajích se měly objevit nové epokcipitály. Pokud se to ovšem skutečně dělo, tak bychom neměli ve fosilním záznamu vidět jen "normální triceratopse" (5 až 7 epiosifikací, bez temenních oken, kratší kulatý límec) a "normální torosaury" (10 a víc epiosifikací, velká temenní okna, dlouhý a hranatý límec), ale i něco mezi: např. triceratopse s malými temenními okny nebo 8 či 9 epiosifikacemi. Podle Scannelly a Hornera (2010, 2011 – viz výše) představuje takovou přechodnou formu jednak Nedoceratops, jednak několik exemplářů triceratopse, v jejichž límci jsou patrné prohlubně v těch místech, kde u "torosaurů" vidíme parietální okna (MOR 2946, AMNH 970). Celé situaci příliš nenapomáhá fakt, že límce jsou často právě v místě těchto prohlubní ulomené (zřejmě kvůli tomu, jak je kost v těchto místech tenká) a ani Farkeho alternativní interpretace kritizovaná výše.
Osobně považuji rozeznání těchto prohlubní za jeden z nejelegantnějších a nejintuitivnějších důkazů, jaké mohli Scannella a Horner pro svou hypotézu přinést, Longrich & Field (2012) jej ale trhají na kusy. Na základě pozorování založených především na exempláři YPM 1823, který Scannella & Horner (2010) citovali jako typickou ukázku daného jevu, konstatují, že prohlubně se tvarem ani pozicí s temenními okny "torosaura" neshodují. Prohlubně na límci triceratopse zasahují na kost šupinovou, okna u "torosaura" jsou celá umístěna výhradně na kosti temenní; autoři si všímají i toho, že za prohlubněmi je límec triceratopse až o 1 cm tlustší než ten "torosaurův". O Longrichově a Fieldsově interpretaci malých oken na límci nedoceratopse už byla řeč: díry opět zasahují do obou skvamosálů a okno v nepoškozené pravé polovině lebky má podivný okraj, nasvědčující tomu, že jde o důsledek zranění či nemoci.
Žádná přechodná fáze nepřeklenuje ani rozdíly v počtu epokcipitálů. Ačkoli Scannella & Horner (2010) navrhli, že 3 až 5 chybějících elementů mohlo triceratopsovi v průběhu života dorůst, Longrich a Field poznamenávají, že není jasné, jak by k tomu mohlo dojít, jelikož tyto přídavné osifikace v průběhu života prostě přirostou k okraji límce a nenechají mezi sebou žádné místo, kam by se mohla vetřít nějaká další. Scannella & Horner (2011; viz výše) si to sice uvědomili a navrhli, že by se epiparietály mohly rozdělit vedví, nepřišli ale s žádným důkazem, že by se něco takového skutečně dělo (připomeňme, že u MOR 2975 se jednalo o episkvamosál, ne o epiparietál).
Posledním výrazným lebečním znakem, který synonymizaci "torosaura" s triceratopsem znesnadňuje, je tvar kostí šupinových. Ty triceratopsovy jsou na horní straně vyduté a mají zesílený vnější okraj, zatímco u "torosaura" jsou ploché a vydutý je naopak jejich vnitřní okraj. Opět neexistují žádné formy, jejichž skvamosály by zaujímaly přechodný tvar mezi oběma extrémy. Scannella & Horner (2010) sice poukázali na to, že rozměry těchto kostí u obou taxonů sdílí stejný poměr délky k relativnímu prodloužení, Longrich & Field (2012) to ale kritizují – jejich graf nezobrazoval vztah mezi dvěma nezávislými veličinami, ale mezi délkou a poměrem délky k šířce; navíc zmiňují dva exempláře "torosaura", jejichž skvamosály jsou protáhlejší, než by z modelu Scannelly a Hornera šlo očekávat.
2.4 Závěr
I přes velice skromné množství subadultních "torosaurů" (s naprostou jistotou v podstatě opět omezených na kontroverzní YPM 1831) autoři nedokázali potvrdit předpověď, podle níž by měly kostry "torosaurů" konzistentně vykazovat větší míru dospělosti než ty triceratopsí. Značný počet triceratopsů totiž jeví jasné známky osteologické vyspělosti, často v kombinaci s malými tělesnými rozměry. Ty vedly Scannellu a Hornera k závěru, že srůsty nejsou dobrým indikátorem věku zvířete; Longrich a Field podporují interpretaci přesně opačnou, totiž že spolehlivým indikátorem není velikost a že různí jedinci přestávali růst při podstatně odlišných rozměrech, snad kvůli pohlavnímu dimorfizmu. Nezanedbatelnou ránu dostal i argument o přechodné morfologii: měl-li Triceratops v průběhu života dospět k "torosauří" morfologii, musel by se nejdřív zbavit 5 až 10 mm tlusté vrstvy kosti ze zadního okraje límce. Longrich & Field (2012) zcela nevylučují, že se nějaké přechodné formy mezi triceratopsí a "torosauří" morfologií ještě objeví, podivují se ale, proč ještě žádné nemáme, když je z pozdní křídy známo ceratopsidních lebek tolik.
Na základě všech těchto faktů se autoři kloní ke konzervativnější taxonomii a triceratopse a "torosaura" označují za dva validní, oddělené taxony. Mají-li pravdu, znamená to, že pořadí srůstů lebečních kostí je u rohatých dinosaurů spolehlivým indikátorem relativního věku jedince (Fujiwara & Takakuwa 2011; contra Scannella & Horner 2010, 2011). Většinu ze srůstů pozorovaných u triceratopse a "torosaura" pozorujeme u jiných chasmosaurinů a lze tedy předpokládat, že se odehrávaly v podobném pořadí. Mnoho z nich rovněž můžeme najít u centrosaurinů, druhé větve skupiny Ceratopsidae. Pokud se stejné indikátory relativního stáří vztahují na více taxonů, můžeme toho využít při diagnóze taxonu nového. Na druhou stranu velikost se nezdá být vhodná k rozlišování ani taxonů, ani ontogenetických stadií. Ukazuje se, že znaky typu počtu epokcipitálů jsou schopny spolehlivě rozlišit různé taxony bez ohledu na jejich ontogenetické stadium. Tento závěr by sice nepřežil, kdyby se synonymizace "torosaura" a triceratopse nakonec ukázala být správnou, ale z hlediska toho, co víme o současných zvířatech, dává docela dobrý smysl. Epiosifikace rohatých dinosaurů jsou totiž zřejmě osteodermy vznikající ve spojitosti se šupinami, a počet ani uspořádání šupin se u žijících sauropsidů během života nemění, díky čemuž jsou taky užitečné při rozlišování jednotlivých taxonů od sebe.
Za hlavní přínos své práce ale Longrich a Field považují zformulování první jasně definované metody, použitelnou k testování hypotéz o synonymitě koncových taxonů*. Takovou dinosauří paleontologie potřebuje jako sůl: diverzitu uměle nafukovala nejen tendence viktoriánských paleontologů popsat každý nediagnostický fragment jako nový "rod" a "druh", ale i záměna rozdílů mezi pohlavími či různě starými zvířaty za rozdíly mezi taxony. Řada "taxonů" ve skutečnosti označuje pouhé juvenilní jedince již dříve známých zvířat: pachycefalosaur Ornatotholus je nejspíš juvenilním stegocerasem (Schott et al. 2011) a jako "Procheneosaurus" byla popisována mláďata několika různých taxonů kachnozobých dinosaurů (Dodson 1975). V mnoha dalších případech se o validitu taxonů vedou dlouhé spory: tyrannosauridní trpaslík Nanotyrannus může být jen mládětem T-rexe (a Longrich a Field ho tak skutečně citují), zároveň ale existují indicie svědčící o opaku (Currie et al. 2003; Larson 2008; Tsuihiji et al. 2011); podobné spory se v současnosti vedou o dalšího malého tyrannosaurida, raptorexe (Fowler et al. 2011 contra Tsuihiji et al. 2011). Kontroverzí se možná dočkáme ještě i v případě drakorexe, na jehož totožnosti s pachycefalosaurem se přitom shodlo vícero nezávislých datasetů (Horner & Goodwin 2009; Longrich et al. 2010), a zřejmě se nevyhne ani synonymitě taxonů Homalocephale a Prenocephale, kterou zatím spíš opatrně navrhli Longrich et al. (2010).
Longrich a Field souhlasí se Scannellou a Hornerem v tom, že při popisu nových taxonů je třeba dávat pozor na ontogenetickou proměnlivost. Podle nich lze odlišné morfologie prohlásit za ontogenetická stadia jednoho taxonu právě tehdy, když 1) se jejich výskyt překrývá v čase i geografii, 2) mezi nimi existují přechodné formy a 3) se exempláře s jednou morfologií na základě nezávislých kritérií zdají být konzistentně starší a "dospělejší" než exempláře vykazující tu druhou. Aplikace těchto kritérií na případ Triceratops/"Torosaurus" podporuje odlišení obou taxonů. Jisté však je, že při budoucím studiu diverzity fosilních dinosaurů bude pochopení ontogenetické proměnlivosti zcela zásadní.
*Pojem "koncový (terminální) taxon" je prodloužením metafory fylogenetického stromu: takto označené skupiny zvířat stojí na koncích jeho větví, v nichž už o žádném dalším dělení na ještě drobnější větvičky nevíme. V úrovňovém názvosloví mohou mít takovéto skupiny taxonomickou kategorii "druhu" (fylogenetický druhový koncept), ale může se jednat i o "rody" (běžné u druhohorních dinosaurů) nebo alopatrické "poddruhy" šířeji chápaného "druhu", jaký produkuje třeba biologický koncept.
3. Budoucnost?
Jsem docela zvědavý, kam nás v taxonomickém konfliktu, který nedávno oslavil třetí narozeniny, dostane další vývoj. Na DML se objevily nářky nad tím, že jeden dataset se dá použít jak k obhajobě, tak vyvrácení "torosaurovy" validity podle toho, kdo ho zrovna analyzuje. Jaime Headden je podstatně optimističtější a (právem) se těší z toho, že velká část debaty se odehrává v žurnálu PLoS ONE, který nejenže poskytuje open-access a přijímá fotografie v mimořádně vysokém rozlišení, ale umožňuje pružnější reakce na publikovaný výzkum díky systému komentářů – zdá se, že i publikační model založený na recenzním řízení se v něčem přiučil od blogů.
Není bez zajímavosti, že studie Longriche a Fielda už obdržela jeden takový komentář od Andrewa Farkeho (autora výše mnohokrát citované studie o nedoceratopsovi), který oba výzkumníky staví do obtížné situace. Souhlasí totiž s jejich závěry, avšak metody, kterým k nim došli, považuje za liché. Do své shlukové analýzy ontogenetických znaků totiž Longrich a Field nezahrnuli počty epiosifikací, tvar temenních kostí ani přítomnost parietálních oken, čímž selhali v testování Scannellova a Hornerova tvrzení, že tyto znaky konzistentně charakterizují dospělejší jedince. Dalo by se dokonce říct, že jejich analýza je vynecháním těchto znaků předpojatá, což pochopitelně obhájcům "torosaurovy" validity zrovna nepomůže. Podle Farkeho pořadí srůstu spolehlivým ontogenetickým indikátorem je, ale analýza Longriche a Fielda není schopna rozhodnout o tom, jakým způsobem jej interpretovat nejlépe.
Zdroje:
V úvodu své studie autoři upozorňují, že pokud by "Torosaurus" a Triceratops skutečně byli různé růstové fáze jediného taxonu, mělo by to dalekosáhlý dopad na dinosauří taxonomii. Bylo by nutné zcela přehodnotit diverzitu rohatých dinosaurů, kde se v současnosti nové taxony rozeznávají právě na základě těch rozdílů, které Scannella a Horner označují za ontogeneticky proměnlivé. Už dnes jsou paleontologové při popisování nových taxonů daleko opatrnější a konzervativnější, než tomu bylo v minulosti (Benton 2008a, b), a synonymizace taxonů zdánlivě tak rozdílných, jako jsou "Torosaurus", Triceratops a Nedoceratops by tento trend jistě ještě posílila. Hypotéza o synonymitě s sebou zároveň nese spoustu testovatelných predikcí, díky čemuž v tomto případě splitting a lumping není otázkou vkusu, ale výzkumu. Longrich a Field formulují následující podmínky, které musí hypotéza o synonymitě splnit: 1) výskyt triceratopse i "torosaura" se musí časově i geograficky překrývat, 2) exempláře obou taxonů musí konzistentně patřit odlišným vývojovým stadiím a 3) musejí existovat nějaké přechodné formy.
2.1 Stratigrafie a biogeografie
Zde hypotéza Scannelly a Hornera na žádné problémy nenaráží. Oba taxony jsou známy ze stejného období (pozdního maastrichtu, tj. nejzazší svrchní křídy) stejné oblasti (západu Severní Ameriky): kostry "torosaura" a triceratopse jsou nalézány ve stejných souvrstvích od Saskatchewanu po Colorado a tam, kde nalézáme jen jednoho z nich (triceratopse v kanadské Albertě, "torosaura" na americkém jihozápadě), klidně může jít o pouhý artefakt nedostatečného fosilního záznamu.

Geografický výskyt triceratopse a "torosaura" vykazuje značnou míru shody. (1) souvrství Scollard, Alberta; (2) souvrství Frenchman, Saskatchewan; (3) souvrství Hell Creek, Montana; (4) souvrství Hell Creek, Severní Dakota; (5) souvrství Hell Creek, Jižní Dakota; (6) souvrství Lance, Wyoming; (7) souvrství Denver, Colorado; (8) souvrství North Horn, Utah; (9) souvrství Javelina; Texas. (Zdroj: Longrich & Field 2012: Figure 2)
2.2 Relativní věk
Je-li "Torosaurus" starý Triceratops, pak by morfologie obou taxonů měla být omezena na určitý věkový interval: neměli bychom nalézat ani příliš mladé "torosaury", ani příliš staré triceratopse. Scannella & Horner (2010) tuto předpověď otestovali histologickým rozborem zaočnicových rohů, Horner & Lamm (2011) pak tutéž metodu aplikovali na temenní kosti. První studie odhalila, že u "torosaura" došlo k rozsáhlejší kostní remodelaci (tj. odbourávání staré kostní tkáně a jejímu nahrazení novou tkání) než u různých exemplářů triceratopse, což si autoři vyložili jako důkaz o jeho větším stáří. Obě studie naneštěstí od "torosaura" prozkoumaly jediný exemplář, takže jejich závěry jsou statisticky naprosto bezcenné. Je také pochybné, že zrovna míra remodelace kosti by měla spolehlivě indikovat dosažené ontogenetické stadium: Lanyon et al. (1982) a O'Connor & Lanyon (1982) totiž experimentem ukázali, že kost může projít přídavnou remodelací, pokud změníme způsob jejího namáhání. Tempo obměňování kostní tkáně se tedy mění podle toho, jak se mění zátěž, kterou kost prodělává, a liší se nejen kost od kosti, ale dokonce i v rámci kosti jediné podle toho, které místo je namáháno nejvíc.
Longrich & Field (2012) pro svou studii vzorek prozkoumaných "torosaurů" podstatně rozšířili, aby zjistili, zda jsou takto označované formy skutečně konzistentně starší než trcieratopsové. Z výše uvedených důvodů k tomu ovšem nevyužívají Scannellovy a Hornerovy histologické rozbory kostěného límce. Histologie dlouhých kostí, která se pro zjišťování stáří zvířete v době smrti obvykle používá, by byla daleko vhodnější; naneštěstí ale není aplikovatelná, neboť většinu exemplářů "torosaura" známe jen z izolované lebky. Nezbývá tedy, než se pokusit odvodit věk jejich materiálů z údajů nashromážděných od kompletnějších rohatých dinosaurů. Je např. známo, že v průběhu stárnutí těchto zvířat prodělávaly jejich lebky řadu morfologických změn: límec se prodlužoval, zaočnicové rohy se protahovaly a zakřivovaly, povrch kosti se měnil z rýhovaného na drsný a sukovitý. Kosti obklopující očnici srostly s nasály a frontály, rostrální kost (jedinečná vývojová novinka rohatých dinosaurů) přirostla ke kosti předčelistní a ta zase ke kosti nosní. Kosti dermálního původu, tvořící rohy nebo okraje kostěného límce, přirostly k příslušným kostem lebky; srostly i některé elementy mozkovny. To má svůj význam, protože u dnešních savců se takové lebeční srůsty odehrávají v typické sekvenci, z níž lze odvodit stáří zvířete (Wang et al. 2006; Sanchez-Villagra 2010). Konkrétně se zdá, že jakmile jsou jednou fúze kostí završeny a veškeré švy uzavřeny, zvíře dosáhlo plné skeletální dospělosti a růst může už jen velice omezeně. Takový indikátor už byl jednou využit při studiu růstových sérií triceratopse (Fujiwara & Takakuwa 2011) a není tedy důvod, proč by se nemohl uplatnit i u "torosaura".
Autoři proto vybrali 24 ontogeneticky proměnlivých osteologických znaků a podívali se, jaké rozdíly vykazují napříč 36 různými exempláři rohatých dinosaurů. Ty pocházejí od taxonů Triceratops horridus, Triceratops prorsus, "Torosaurus latus", "Torosaurus utahensis" a Tatankaceratops sacrisonorum. Už jen za tímto základním pracovním roztříděním stojí obrovské množství kontroverze: pomineme-li samotné oddělování "torosaura" a triceratopse (prováděné z větší části na základě právě těch kontroverzních znaků, jejichž ontogenetická proměnlivost se teprve má testovat), status "T. utahensis" je velice nejistý a Tatankaceratops je až zarážejícně podobný T. prorsus – až na to, že podivným způsobem míchá znaky typické pro mladá a dospělá zvířata. Podle Longriche (2010) jde buď o exemplář prorsa, který přestal růst ještě před dosažením běžné velikosti, nebo nový, trpasličí subtaxon triceratopse. Zvláštní zmínku si zaslouží Nedoceratops, který je prý ve skutečnosti jen poškozeným exemplářem T. horridus: díry v límci jsou patologické a nejde o pravá temenní okna (vzhledem k jejich nepravidelnému tvaru), zaočnicový roh kolmý na zubní řadu pak lze údajně nalézt jen na jedné straně lebky a jde tak jen o tafonomický artefakt. Zdá se tedy, že už tu máme minimálně tři konfliktní interpretace USNM 2412: Longrich s Fieldem a Scannella s Hornerem se shodnou na tom, že jde o triceratopse (contra Farke 2011), ne už ale na tom, zda je jeho divná lebka jedinečnou ukázkou "přechodné morfologie", nebo jen trochu rozmáčknutá. Ať tak či onak, nashromážděná ontogeneticky informativní data autoři podrobili rozboru metodou maximální parsimonie, která vzhledem k velkému množství chybějících dat a malé variabilitě našla extrémně velké množství optimálních stromů. Autoři ji nechali běžet jen do 250 000 stromů, z nichž byl následně vytvořen striktní konsenzus přetištěný níže (tj. strom obsahující jen ty shluky, které byly přítomné na každém stromě).

Shluková analýza jedinců náležejících k různým taxonům chasmosaurinních rohatých dinosaurů, vytvořená metodou nejvyšší parsimonie. Výsledný diagram by neměl být čten jako běžný fylogenetický strom; má se číst lineárně, divergence nejsou podstatné. Seznamy na jednotlivých "větvích" zachycují, jaké morfologické změny provázely stárnutí zvířete. Původní analýza dokázala rozřešit stáří zde nezahrnutých exemplářů MNHN 1912.20, YPM 1830, YPM 1828, USNM 5740 (dospělci) a USNM 15583 (subadultní jedinec), které ovšem svými chybějícími daty způsobily ztrátu rozlišení v jiných místech stromu. Z dalšího běhu analýzy, jehož výsledky ilustruje tento obrázek, je proto autoři vyřadili. Hvězdičky označují morfologické změny, jejichž ontogenetický význam je nejistý v důsledku nedostatečných nebo konfliktních údajů. (Zdroj: Longrich & Field 2012: Figure 5)
Výsledný strom v tomto případě nereprezentuje průběh fylogeneze, nýbrž ontogeneze: jedinci posazení na jeho bázi jsou nejmladší, ti na "vršku" zase nejstarší. Model to samozřejmě není ideální: ačkoli je datová matrice malá, vyskytly se v ní znakové konflikty a problémem jsou i již zmiňovaná chybějící data. Přesto jsou na výsledném stromě rozeznatelné tři odlišné fáze, které Longriech a Field ztotožňují s mláďaty, subadultními jedinci a dospělci. V první fázi se protáhly a rozšířily temenní kosti a zaočnicové rohy se prodloužily, zmohutněly a zakřivily. V druhé fázi spolu navzájem srostly kosti nosní, slzné, předčelní, čelní a zaočnicové a několik kostí mozkovny; textura tvářových kostí a límce začala pomalu hrubnout. Překvapivé je, že v této fázi najdeme největší zdokumentovaná zvířata vůbec. Patří sem totiž i již zmiňovaný "subadultní torosaur" YPM 1831, vůbec největší zástupce svého taxonu, nebo lebka triceratopse s katalogovým označením YPM 1821, která s délkou 101 cm o plných 15 cm přesahuje lebky některých dospělců. (Longrich a Field ujišťují, že podobné nesrovnalosti mezi tělesnými rozměry a dosaženým stupněm ontogeneze panují i u triceratopse.) Konečně během třetí fáze přirostly epokcipitály k límci, epijugály ke kosti jařmové a nosní kost srostla s premaxillou a rostrálem. Autoři také zaznamenali jisté individuální odchylky: změna textury zřejmě u některých jedinců nastala později než u jiných. Matoucí znakové konflikty nastávají i jinde: YPM 1831 nemá srostlý týlní hrbol – na rozdíl od zvířat, která se podle ostatních znaků zdají být mladší – a Tatankaceratops směšuje znaky juvenilních a nedospělých jedinců.
V souvislosti s vyvracením Scannellovy a Hornerovy hypotézy je ale nejzajímavější to, kde se umístilo šest prozkoumaných "torosaurů". Tři z nich se zařadili do kategorie dospělců, minimálně jeden z nich se však jeví mladší, než mnohé exempláře triceratopse. Dva další (včetně tolik problematického YPM 1831) stojí na rozmezí kategorie dospělců a subadultních jedinců; YPM 1831 se nakonec skutečně zdá být spíše subadultní, jak o něm napsal Farke (2011). Důležité ale je, že analýza Longriche a Fielda tento závěr podložila nezávislou evidencí a subadultní status pro YPM 1831 není založen na znacích, které kritizovali Scannella & Horner (2011; viz výše). Místo toho jej podporuje volný epijugál, rostrální kost nepřirostlá k premaxille a již zmiňovaný nesrostlý týlní hrbol; tedy znaky, které u dospělců rohatých dinosaurů nejsou obvyklé. Je pravda, že ani sekvence fúzí lebečních kostí není podle Scannelly a Hornera ontogeneticky informativní; s touto námitkou se však autoři už vypořádali výše. Subadultním se dále zdá být i "Torosaurus" utahensis. Analýza naopak odhalila poměrně velké množství (10) dospělých triceratopsů, což ostře kontrastuje s výsledky Scannelly a Hornera (2010), kteří nebyli schopni nalézt ani jednoho. Podle Longriche a Fielda je to způsobeno tím, že předchozí dvojice autorů zařadila do své studie jen dva dostatečně velké triceratopsí exempláře. Žádný juvenilní "Torosaurus", který by Scannellovu a Hornerovu hypotézu vyvrátil nejlépe, v analýze identifikován nebyl.

Nedospělé znaky na YPM 1831. Ačkoli je tato lebka s délkou více než 2,6 m vůbec největším známým exemplářem "torosaura", analýza Longriche a Fielda ji odhalila jako subadultního jedince. Tento závěr stojí a padá s tím, nakolik spolehlivá je posloupnost srůstů lebečních elementů při určování stáří: Longrich a Field tvrdí, že je, a načrtávají analogie s dnešními savci; Scannella a Horner tvrdí, že není, a poukazují na rozpory mezi tělesnými rozměry a počtem dokončených fúzí, jichž je YPM 1831 sama dobrým příkladem. Na obrázku (A) je zachycen epijugál – epiosifikace přirůstající ke kosti jařmové, u tohoto exempláře však stále samostatná – z předního (A1), vnějšího bočního (A2) a vnitřního (A3) pohledu. V (B) je zachycen rostrál – kost unikátní pro ptakopánvé dinosaury, tvořící horní polovinu ceratopsího zobáku – stále oddělený od maxily, a to seshora (B1) a zespoda (B2). (C) týlní hrbol je tvořen ještě nesrostlými exokcipitály (bočními týlními kostmi) a baziokcipitálem (spodní týlní kostí). (D) zaoblený okraj temenní kosti s ještě nepřirostlými epiosifikacemi. (E) dlouhozrnná textura na povrchu temenní kosti. Proti ontogenetickému významu posledních dvou znaků samozřejmě argumentovali Scannella & Horner (2011; viz výše). Pruh měřítka odpovídá 5 cm na obrázcích A, B a 2 cm na E. (Zdroj: Longrich & Field 2012: Figure 7)
2.3 Přechodné formy
Pokud mají Scannella a Horner pravdu a triceratopsi svůj život končili s "torosauří" morfologií lebky, musela jejich lebka v průběhu života absolvovat výrazné morfologické změny: límec se měl protáhnout a změnit tvar, resorpcí kostní tkáně měla vzniknout temenní okna a na jeho okrajích se měly objevit nové epokcipitály. Pokud se to ovšem skutečně dělo, tak bychom neměli ve fosilním záznamu vidět jen "normální triceratopse" (5 až 7 epiosifikací, bez temenních oken, kratší kulatý límec) a "normální torosaury" (10 a víc epiosifikací, velká temenní okna, dlouhý a hranatý límec), ale i něco mezi: např. triceratopse s malými temenními okny nebo 8 či 9 epiosifikacemi. Podle Scannelly a Hornera (2010, 2011 – viz výše) představuje takovou přechodnou formu jednak Nedoceratops, jednak několik exemplářů triceratopse, v jejichž límci jsou patrné prohlubně v těch místech, kde u "torosaurů" vidíme parietální okna (MOR 2946, AMNH 970). Celé situaci příliš nenapomáhá fakt, že límce jsou často právě v místě těchto prohlubní ulomené (zřejmě kvůli tomu, jak je kost v těchto místech tenká) a ani Farkeho alternativní interpretace kritizovaná výše.
Osobně považuji rozeznání těchto prohlubní za jeden z nejelegantnějších a nejintuitivnějších důkazů, jaké mohli Scannella a Horner pro svou hypotézu přinést, Longrich & Field (2012) jej ale trhají na kusy. Na základě pozorování založených především na exempláři YPM 1823, který Scannella & Horner (2010) citovali jako typickou ukázku daného jevu, konstatují, že prohlubně se tvarem ani pozicí s temenními okny "torosaura" neshodují. Prohlubně na límci triceratopse zasahují na kost šupinovou, okna u "torosaura" jsou celá umístěna výhradně na kosti temenní; autoři si všímají i toho, že za prohlubněmi je límec triceratopse až o 1 cm tlustší než ten "torosaurův". O Longrichově a Fieldsově interpretaci malých oken na límci nedoceratopse už byla řeč: díry opět zasahují do obou skvamosálů a okno v nepoškozené pravé polovině lebky má podivný okraj, nasvědčující tomu, že jde o důsledek zranění či nemoci.
Žádná přechodná fáze nepřeklenuje ani rozdíly v počtu epokcipitálů. Ačkoli Scannella & Horner (2010) navrhli, že 3 až 5 chybějících elementů mohlo triceratopsovi v průběhu života dorůst, Longrich a Field poznamenávají, že není jasné, jak by k tomu mohlo dojít, jelikož tyto přídavné osifikace v průběhu života prostě přirostou k okraji límce a nenechají mezi sebou žádné místo, kam by se mohla vetřít nějaká další. Scannella & Horner (2011; viz výše) si to sice uvědomili a navrhli, že by se epiparietály mohly rozdělit vedví, nepřišli ale s žádným důkazem, že by se něco takového skutečně dělo (připomeňme, že u MOR 2975 se jednalo o episkvamosál, ne o epiparietál).
Posledním výrazným lebečním znakem, který synonymizaci "torosaura" s triceratopsem znesnadňuje, je tvar kostí šupinových. Ty triceratopsovy jsou na horní straně vyduté a mají zesílený vnější okraj, zatímco u "torosaura" jsou ploché a vydutý je naopak jejich vnitřní okraj. Opět neexistují žádné formy, jejichž skvamosály by zaujímaly přechodný tvar mezi oběma extrémy. Scannella & Horner (2010) sice poukázali na to, že rozměry těchto kostí u obou taxonů sdílí stejný poměr délky k relativnímu prodloužení, Longrich & Field (2012) to ale kritizují – jejich graf nezobrazoval vztah mezi dvěma nezávislými veličinami, ale mezi délkou a poměrem délky k šířce; navíc zmiňují dva exempláře "torosaura", jejichž skvamosály jsou protáhlejší, než by z modelu Scannelly a Hornera šlo očekávat.
2.4 Závěr
I přes velice skromné množství subadultních "torosaurů" (s naprostou jistotou v podstatě opět omezených na kontroverzní YPM 1831) autoři nedokázali potvrdit předpověď, podle níž by měly kostry "torosaurů" konzistentně vykazovat větší míru dospělosti než ty triceratopsí. Značný počet triceratopsů totiž jeví jasné známky osteologické vyspělosti, často v kombinaci s malými tělesnými rozměry. Ty vedly Scannellu a Hornera k závěru, že srůsty nejsou dobrým indikátorem věku zvířete; Longrich a Field podporují interpretaci přesně opačnou, totiž že spolehlivým indikátorem není velikost a že různí jedinci přestávali růst při podstatně odlišných rozměrech, snad kvůli pohlavnímu dimorfizmu. Nezanedbatelnou ránu dostal i argument o přechodné morfologii: měl-li Triceratops v průběhu života dospět k "torosauří" morfologii, musel by se nejdřív zbavit 5 až 10 mm tlusté vrstvy kosti ze zadního okraje límce. Longrich & Field (2012) zcela nevylučují, že se nějaké přechodné formy mezi triceratopsí a "torosauří" morfologií ještě objeví, podivují se ale, proč ještě žádné nemáme, když je z pozdní křídy známo ceratopsidních lebek tolik.
Na základě všech těchto faktů se autoři kloní ke konzervativnější taxonomii a triceratopse a "torosaura" označují za dva validní, oddělené taxony. Mají-li pravdu, znamená to, že pořadí srůstů lebečních kostí je u rohatých dinosaurů spolehlivým indikátorem relativního věku jedince (Fujiwara & Takakuwa 2011; contra Scannella & Horner 2010, 2011). Většinu ze srůstů pozorovaných u triceratopse a "torosaura" pozorujeme u jiných chasmosaurinů a lze tedy předpokládat, že se odehrávaly v podobném pořadí. Mnoho z nich rovněž můžeme najít u centrosaurinů, druhé větve skupiny Ceratopsidae. Pokud se stejné indikátory relativního stáří vztahují na více taxonů, můžeme toho využít při diagnóze taxonu nového. Na druhou stranu velikost se nezdá být vhodná k rozlišování ani taxonů, ani ontogenetických stadií. Ukazuje se, že znaky typu počtu epokcipitálů jsou schopny spolehlivě rozlišit různé taxony bez ohledu na jejich ontogenetické stadium. Tento závěr by sice nepřežil, kdyby se synonymizace "torosaura" a triceratopse nakonec ukázala být správnou, ale z hlediska toho, co víme o současných zvířatech, dává docela dobrý smysl. Epiosifikace rohatých dinosaurů jsou totiž zřejmě osteodermy vznikající ve spojitosti se šupinami, a počet ani uspořádání šupin se u žijících sauropsidů během života nemění, díky čemuž jsou taky užitečné při rozlišování jednotlivých taxonů od sebe.
Za hlavní přínos své práce ale Longrich a Field považují zformulování první jasně definované metody, použitelnou k testování hypotéz o synonymitě koncových taxonů*. Takovou dinosauří paleontologie potřebuje jako sůl: diverzitu uměle nafukovala nejen tendence viktoriánských paleontologů popsat každý nediagnostický fragment jako nový "rod" a "druh", ale i záměna rozdílů mezi pohlavími či různě starými zvířaty za rozdíly mezi taxony. Řada "taxonů" ve skutečnosti označuje pouhé juvenilní jedince již dříve známých zvířat: pachycefalosaur Ornatotholus je nejspíš juvenilním stegocerasem (Schott et al. 2011) a jako "Procheneosaurus" byla popisována mláďata několika různých taxonů kachnozobých dinosaurů (Dodson 1975). V mnoha dalších případech se o validitu taxonů vedou dlouhé spory: tyrannosauridní trpaslík Nanotyrannus může být jen mládětem T-rexe (a Longrich a Field ho tak skutečně citují), zároveň ale existují indicie svědčící o opaku (Currie et al. 2003; Larson 2008; Tsuihiji et al. 2011); podobné spory se v současnosti vedou o dalšího malého tyrannosaurida, raptorexe (Fowler et al. 2011 contra Tsuihiji et al. 2011). Kontroverzí se možná dočkáme ještě i v případě drakorexe, na jehož totožnosti s pachycefalosaurem se přitom shodlo vícero nezávislých datasetů (Horner & Goodwin 2009; Longrich et al. 2010), a zřejmě se nevyhne ani synonymitě taxonů Homalocephale a Prenocephale, kterou zatím spíš opatrně navrhli Longrich et al. (2010).
Longrich a Field souhlasí se Scannellou a Hornerem v tom, že při popisu nových taxonů je třeba dávat pozor na ontogenetickou proměnlivost. Podle nich lze odlišné morfologie prohlásit za ontogenetická stadia jednoho taxonu právě tehdy, když 1) se jejich výskyt překrývá v čase i geografii, 2) mezi nimi existují přechodné formy a 3) se exempláře s jednou morfologií na základě nezávislých kritérií zdají být konzistentně starší a "dospělejší" než exempláře vykazující tu druhou. Aplikace těchto kritérií na případ Triceratops/"Torosaurus" podporuje odlišení obou taxonů. Jisté však je, že při budoucím studiu diverzity fosilních dinosaurů bude pochopení ontogenetické proměnlivosti zcela zásadní.
*Pojem "koncový (terminální) taxon" je prodloužením metafory fylogenetického stromu: takto označené skupiny zvířat stojí na koncích jeho větví, v nichž už o žádném dalším dělení na ještě drobnější větvičky nevíme. V úrovňovém názvosloví mohou mít takovéto skupiny taxonomickou kategorii "druhu" (fylogenetický druhový koncept), ale může se jednat i o "rody" (běžné u druhohorních dinosaurů) nebo alopatrické "poddruhy" šířeji chápaného "druhu", jaký produkuje třeba biologický koncept.
3. Budoucnost?
Jsem docela zvědavý, kam nás v taxonomickém konfliktu, který nedávno oslavil třetí narozeniny, dostane další vývoj. Na DML se objevily nářky nad tím, že jeden dataset se dá použít jak k obhajobě, tak vyvrácení "torosaurovy" validity podle toho, kdo ho zrovna analyzuje. Jaime Headden je podstatně optimističtější a (právem) se těší z toho, že velká část debaty se odehrává v žurnálu PLoS ONE, který nejenže poskytuje open-access a přijímá fotografie v mimořádně vysokém rozlišení, ale umožňuje pružnější reakce na publikovaný výzkum díky systému komentářů – zdá se, že i publikační model založený na recenzním řízení se v něčem přiučil od blogů.
Není bez zajímavosti, že studie Longriche a Fielda už obdržela jeden takový komentář od Andrewa Farkeho (autora výše mnohokrát citované studie o nedoceratopsovi), který oba výzkumníky staví do obtížné situace. Souhlasí totiž s jejich závěry, avšak metody, kterým k nim došli, považuje za liché. Do své shlukové analýzy ontogenetických znaků totiž Longrich a Field nezahrnuli počty epiosifikací, tvar temenních kostí ani přítomnost parietálních oken, čímž selhali v testování Scannellova a Hornerova tvrzení, že tyto znaky konzistentně charakterizují dospělejší jedince. Dalo by se dokonce říct, že jejich analýza je vynecháním těchto znaků předpojatá, což pochopitelně obhájcům "torosaurovy" validity zrovna nepomůže. Podle Farkeho pořadí srůstu spolehlivým ontogenetickým indikátorem je, ale analýza Longriche a Fielda není schopna rozhodnout o tom, jakým způsobem jej interpretovat nejlépe.
Zdroje:
- http://www.plosone.org/annotation/listThread.action?inReplyTo...
- Archibald DJ 1996 Dinosaur extinction and the end of an era: what the fossils say. New York: Columbia Univ Press. 237 p
- Benton MJ 2008a How to find a dinosaur, and the role of synonymy in biodiversity studies. Paleobiol 34(4): 516–33
- Benton MJ 2008b Fossil quality and naming dinosaurs. Biol Lett 4(6): 729–32
- Brown CM, Russell AP, Ryan MJ 2009 Pattern and transition of surficial bone texture of the centrosaurine frill and their ontogenetic and taxonomic implications. J Vert Paleont 29(1): 132–41
- Campione NE, Evans DC 2011 Cranial growth and variation in Edmontosaurs (Dinosauria: Hadrosauridae): implications for Latest Cretaceous megaherbivore diversity in North America. PLoS ONE 6(9): e25186
- Cobabe EA, Fastovsky DE 1987 Ugrosaurus olsoni, a new ceratopsian (Reptilia: Ornithischia) from the Hell Creek Formation of eastern Montana. J Paleont 61(1): 148–54
- Currie PJ, Hurum JH, Sabath K 2003 Skull structure and evolution in tyrannosaurid dinosaurs. Acta Palaeont Pol 48(2): 227–34
- Dodson P 1975 Taxonomic implications of relative growth in lambeosaurine hadrosaurs. Syst Zool 24(1): 37–54
- Dodson P, Forster CA, Sampson SD 2004 Ceratopsidae. 494–513 in Weishampel DB, Dodson P; Osmólska H, eds. The Dinosauria 2nd edition. Berkeley, CA: Univ of California Press
- Farke AA 2011 Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE 6(1): e16196
- Forster CA 1993 Taxonomic validity of the ceratopsid dinosaur Ugrosaurus olsoni(CoBabe and Fastovsky). J Paleont 67(2): 316–8
- Forster CA 1996a Species resolution in Triceratops: cladistic and morphometric approaches. J Vert Paleont 16(2): 259–70
- Forster CA 1996b New information on the skull of Triceratops. J Vert Paleont 16(2): 246–58
- Forster CA, Sereno PC, Evans TW, Rowe T 1993 A complete skull of Chasmosaurus mariscalensis (Dinosauria: Ceratopsidae) from the Aguja Formation (Late Campanian) of West Texas. J Vert Paleont 13(2): 161–70
- Fowler DW, Woodward HN, Freedman EA, Larson PL, Horner JR 2011 Reanalysis of “Raptorex kriegsteini”: A juvenile tyrannosaurid dinosaur from Mongolia. PLoS ONE 6(6): e21376
- Fujiwara S-i, Takakuwa Y 2011 A sub-adult growth stage indicated in the degree of suture co-ossification in Triceratops. Bull Gunma Mus Nat Hist 15: 1–17
- Godfrey SJ, Holmes R 1995 Cranial morphology and systematics of Chasmosaurus (Dinosauria: Ceratopsidae) from the Upper Cretaceous of Western Canada. J Vert Paleont 15(4): 726–42
- Horner JR, Goodwin MB 2006 Major cranial changes during Triceratops ontogeny. Proc R Soc B 273(1602): 2757–61
- Horner JR, Goodwin MB 2008 Ontogeny of cranial epi-ossifications in Triceratops. J Vert Paleont 28(1): 134–44
- Horner JR, Goodwin MB 2009 Extreme cranial ontogeny in the Upper Cretaceous dinosaur Pachycephalosaurus. PLoS ONE 4(10): e27626
- Horner JR, Goodwin MB, Myrhvold N 2011 Dinosaur census reveals abundant Tyrannosaurus and rare ontogenetic stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA. PLoS ONE 6(2): e16574
- Horner JR, Lamm E-T 2011 Ontogeny of the parietal frill of Triceratops: a preliminary histological analysis. Comptes Rendus Palevol 10(5–6): 439–452
- Lanyon LE, Goodship AE, Pye CJ, MacFie JH 1982 Mechanically adaptive bone remodelling. J Biomech 15(3): 141–54
- Larson P 2008 Variation and sexual dimorphism in Tyrannosaurus rex. 102–28 in Larson P, Carpenter K, eds. Tyrannosaurus rex, the tyrant king. Bloomington, IN: Indiana Univ Press
- Lehman TM 1990 The ceratopsian subfamily chasmosaurinae: sexual dimorphism and systematics. 211–29 in Carpenter K, Currie PJ, eds. Dinosaur systematics: approaches and perspectives. New York: Cambridge Univ Press
- Lehman TM 1998 A gigantic skull and skeleton of the horned dinosaur Pentaceratops sternbergi from New Mexico. J Paleont 72(5): 894–906
- Longrich NR 2010 Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretac Res 32: 264–76
- Longrich NR, Field DJ 2012 Torosaurus is not Triceratops: Ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy. PLoS ONE 7(2): e32623
- Longrich NR, Sankey JT, Tanke DH 2010 Texacephale langstoni, a new genus of pachycephalosaurid (Dinosauria: Ornithischia) from the upper Campanian Aguja Formation, southern Texas, USA. Cretac Res 31(2): 274–84
- Lull RS 1905 Restoration of the horned dinosaur Diceratops. Am J Sci 20: 420–2
- Lull RS 1933 A revision of the Ceratopsia or horned dinosaurs. Yale Peabody Mus Mem 3: 1–175
- Mateus O 2008 Two ornithischian dinosaurs renamed: Microceratops Bohlin, 1953 and Diceratops Lull, 1905. J Paleont 82(2): 423
- O’Connor JA, Lanyon LE 1982 Influence of strain rate on adaptive bone remodelling. J Biomech 15(10): 767–81
- Ostrom JH, Wellnhofer P 1986 The Munich specimen of Triceratops with a revision of the genus. Zitteliana 14: 111–58
- Sampson SD, Ryan MJ, Tanke DH 1997 Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taxonomic and behavioral implications. Zool J Linn Soc 121(3): 293–337
- Sanchez-Villagra MR 2010 Suture closure as a paradigm to study late growth in recent and fossil mammals: a case study with giant deer and dwarf deer skulls. J Vert Paleont 30(6): 1895–8
- Scannella JB, Horner JR 2010 Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny. J Vert Paleont 30(4): 1157–68
- Scannella JB, Horner JR 2011 ‘Nedoceratops’: an example of a transitional morphology. PLoS ONE 6: e28705
- Schott RK, Evans DC, Goodwin MB, Horner JR, Brown CM, Longrich NR 2011 Cranial ontogeny in Stegoceras validum (Dinosauria: Pachycephalosauria): A quantitative model of pachycephalosaur dome growth and variation. PLoS ONE 6(6): e21092
- Tsuihiji T 2010 Reconstructions of the axial muscle insertions in the occipital region of dinosaurs: evaluations of past hypotheses on Marginocephalia and Tyrannosauridae using the Extant Phylogenetic Bracket approach. Anat Rec 293(8): 1360–86
- Tsuihiji T, Watabe M, Tsogtbaatar K, Tsubamoto T, Barsbold R, Suzuki S, Lee AH, Ridgely RC, Kawahara Y, Witmer LM 2011 Cranial osteology of a juvenile specimen of Tarbosaurus bataar (Theropoda, Tyrannosauridae) from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia. J Vert Paleont 31(3): 497—517
- Ukrainsky AS 2007 A new replacement name for Diceratops Lull, 1905 (Reptilia: Ornithischia: Ceratopsidae). Zoosyst Ross 16: 292
- Wang Q, Strait DS, Dechow PC 2006 Fusion patterns of craniofacial sutures in rhesus monkey skulls of known age and sex from Cayo Santiago. Am J Phys Anthropol 131(4): 469–486.










