
PLoS ONE je dinosauřím nadšencům dobře známý: především jako "proof of concept" online žurnál, publikující práce vysoké kvality pod modelem open access, díky němuž si je může zcela zdarma přečíst každý, kdo o ně projeví zájem. Papír je potřeba jen k tomu, aby popisy nových taxonů splnily poněkud zastaralé požadavky ICZN. Autoři publikující v PLoS ONE nejsou limitováni prostorem a mohou tedy každý objev rozebrat tak podrobně, jak jen uznají za vhodné. Stejně tak obrazový materiál může být prezentován v kvalitě, kterou jinde nevidíme, což velmi dobře využijí právě paleontologické studie.
V uplynulých dvou měsících se v PLoS ONE objevilo hned několik zajímavých prací o dinosaurech. Protože spolu opravdu nemají společného víc, než místo publikace, nebudu z nich sestavovat sérii (jako byla např. ta o konfliktu molekul s morfologií). Dva z nich jsou si ovšem tak blízké, že jsem je přeci jen do jednoho článku spojil: v rozmezí dvou dní byly ve stejném periodiku publikovány dvě práce, zabývající se pachycefalosaurem (tlustolebým dinosaurem) stegocerasem. Právě ty se v nynějším článku snažím shrnout.
1. Snively & Theodor (2011) o pachycefalosauřím trkání
Pachycefalosauři jsou malou, avšak velmi výraznou skupinou malých až středně velkých ptakopánvých dinosaurů, podle dnešních hypotéz blízce příbuzných rohatým ceratopsům. Přestože jsou nejlépe známi svou "kupolí" na hlavě, tvořenou extrémně zesílenými čelními a temenními kostmi, ne všichni známí zástupci této skupiny tímto znakem disponují, a to i přesto, že samo jméno Pachycephalosauria znamená doslova "tlustohlaví ještěři.
Historicky byla poměrně oblíbená hypotéza, že pachycefalosauři užívali zesílené lebeční kosti k podobným soubojům, jako dnešní ovce tlustorohé (Ovis canadensis) nebo antilopy chocholatky, které sdílí s pachycefalosaury velmi tlusté, zaoblené kosti čelní. Dinosauří renezance 70. let, v níž paleontologové poprvé po dlouhé době začali o dinosaurech uvažovat jako o aktivních zvířatech, podobných spíše dnešním savcům než krokodýlům, rychle přijala hypotézu o trkání za svou. Sues (1978) např. demonstroval její biomechanickou přijatelnost na plexisklových modelech. Jenže jako tomu bylo i s jinými, často až příliš "heretickými" myšlenkami dinosauří renezance, ani představa pachycefalosaurů svádějících souboje prudkými údery hlavou o hlavu nepřečkala důkladnější rozbor. Zamítla ji histologická studie Goodwina a Hornera (2004), podle níž takovou činnost nedovolovala struktura kupole, v níž se pod povrchovou vrstvou kompaktní kosti skrývají sloupcovité trámečky (trabeculae) kolmé na vnější povrch dómu. Jiné práce ale tento výsledek zpochybnily: Snively & Cox (2008) argumentovali, že pokud by kostěnou kupoli pokrývala dostatečně silná vrstva rohoviny, nemožné by to být nemuselo; a trámcovitá struktura mohla být elastičtější, než si Goodwin a Horner mysleli. Farke (2008) zase s využitím metody konečných prvků ukázal, že lebeční dutiny u kozy domácí (Capra hircus) nezpůsobují snížení mechanické zátěže při nárazu, jak se dosud myslelo – ba právě naopak, modely s kompaktními čelními kostmi vykazovaly menší zátěž, a modely s vyklenutými, tlustými čelními kostmi často dokonce ještě menší. Hypotéza, že lebeční dutiny nebo kosti, které tyto dutiny ohraničují, dovedou absorbovat šok, tak byla jeho daty spíše vyvrácena. Absence vzduchem vyplněných prostor v lebečních kupolích pachycefalosaurů přitom byla v minulosti rovněž použita proti hypotéze o trkání (Fastovsky & Weishampel 2005).
Snively & Theodor (2011) se proto nyní pokoušejí dobrat se z těchto vzájemně rozporných indicií konečné odpovědi. Jejich studie aplikuje CT sken a 3D analýzu metodou konečných prvků na taxony, od nichž trkání hlavami prokazatelně známe (sudokopytníci chocholatka bělobřichá [Cephalophus leucogaster], pižmoň severní [Ovibos moschatus] a žirafa [Giraffa camelopardalis]), a na taxon, který je z trkání podezřelý – pachycefalosaur Stegoceras validum. Jelikož se ale žijící savci mlátí hlavami velmi vynalézavými způsoby, zahrnuli do analýzu taky ovci tlustorohou, která se vyhýbá kontaktu "roh na roh"; vidloroha amerického (Antilocapra americana), který naopak využívá své rohy k přetlačování a povrchem hlavy netrká; a také wapitiho (Cervus canadensis), pekari páskovaného (Tayasu tajacu) a lamu krotkou (Lama glama), kteří nedělají ani jedno.
Autoři dosti originálně nevyužívají data získaná od sudokopytníků k predikcím o stegocerasovi, ale data o stavbě stegocerasovy lebky k predikcím o stavbě lebek sudokopytníků. Konkrétně Snively a Theodor stavějí na tom, že v lebeční kupoli pachycefalosaurů (označované též přesnějším pojmem frontoparietální, který říká, že se skládá z kostí čelních [frontálů] a temenních [parietálů]) najdeme hluboko pod vrstvou kompaktní hutné kosti houbovitou kostní tkáň, a očekávají, že u turovitých bychom měli nalézt stejné uspořádání. Dále testují, zda kombinace lebečních dutin a hutné kosti umožňuje účinnější rozptyl tlaku, zda (a případně jak) by různě tvarované rohovinové polštáře v tomto ohledu pomohly, a zda bude stejný náraz na lebky stegocerase a žijících savců působit stejnou mechanickou námahou. Využívají k tomu algoritmus rekurzivního dělení (recursive partitioning analysis), který je znám především svým uplatněním medicíně: umožňuje stanovit diagnózu na základě známých korelací mezi symptomy a nemocemi. Snively & Theodor (2011) jej využívají přesně analogicky, když chtějí diagnostikovat chování zvířete z jeho morfologických znaků.

CT sken odhalil, že frontoparietální kupole stegocerase se skládá z kosti s vysokou a rovnoměrnou hustotou, větší než je hustota kostěného patra nebo týlu. To částečně souhlasí s daty získanými od žijící chocholatky, kde je vnější povrch "kupole", tvořené vyklenutou částí čelních kostí, také hustší než okolí. U chocholatky ale dosahují srovnatelné hustoty také nosní kosti a boční část kostí temenních. Proximální (povrchu lebky bližší) část kostěných jader rohů je zde daleko hustší než rohovinová vrstva, která jádra obaluje. U pižmoně je tomu naopak: rohovina je hustší než houbovitá kost, z níž je tvořena lebka.
Pokud se ale zaměříme na vnitřek kosti a ne jen její povrch, zjistíme, že mezi dvěma vrstvami velmi husté a kompaktní kosti (histologické zóny 1 a 3 podle Goodwina a Hornera [2004]) se nachází méně hustý prostředek (zóna 2). Svrchní zóna kompaktní kosti je tlustší na špičce kupole než po jejích okrajích. Dutiny, v nichž se za života zvířete nacházely cévy, se sice nacházejí ve vrstvě houbovité kosti, jsou však ohraničeny kostí kompaktní, ačkoli méně hustou, než je ta na povrchu kupole. Tato kompaktní kost tedy formuje trubice, které ústí na vnějším povrchu kupole. Podobně hustá kost rovněž tvoří hluboko uvnitř kupole jakési "proužky" (patrné na obrázku výše na snímcích C a D), které jsou zřejmě pozůstatkem raných stadií růstu kupole (což je téma, které je podrobně rozebráno níže). Rozložení různě hustých kostí u stegocerase kopíruje to, které známe od chocholatky. I tam najdeme hustou, kompaktní vrstvu přilehlou k mozkovně, nad kterou se klene nejprve houbovitá a blízko povrchu opět velice hustá kompaktní kostní tkáň. Velikost a hustota kostních trámečků je u chocholatky rozmanitější než u stegocerase: zatímco u vymřelého pachycefalosaura vybíhají kolmo k vnějšímu povrchu, u žijícího sudokopytníka vytvářejí houbovité mřížoví jakýchsi podpěr. Rovněž u ovce tlustorohé vidíme vrstvy v pořadí hluboká kompaktní, střední houbovitá a povrchová kompaktní. Pro pižmoně toto uspořádání platí v zásadě také (zaměříme-li se na relevantní část lebky, což v tomto případě není vyklenuté čelo, ale zkroucené rohy), chybí mu ale spodní vrstva kompaktní mozkovny obklopující mozkovnu. Ta je místo toho ohraničena houbovitou kostí, která pokračuje skrze většinu délky rohů až po špičku, kde je vystřídána vrstvou kompaktní kosti, překrytou následně ještě rohovinovou pochvou. Tato vrstva je tenčí než u stegocerase. Na rozdíl od tohoto pachycefalosaura také hustota povrchové vrstvy čelních kostí pižmoně nepřevyšuje hustotu nasálů a kostí horní čelisti.
U dalších sudokopytníků se lebeční dutiny, houbovitá kost a kompaktní kost vyskytují v nejrůznějších kombinacích. Žirafa má vrstvu houbovité kosti podstatně tenčí a objem dutin nad mozkem podstatně větší, než ovce tlustorohá. V jejích růžcích (osikonech) nicméně najdeme povrchovou vrstvu velmi husté kompaktní kosti a hluboko položenou vrstvu stejné, avšak o něco řidší kostní tkáně právě tak, jak je tomu u stegocerase. Pekari má nad mozkovnou dutiny, ale ne houbovitou kost, a wapiti, lama i vidloroh postrádají obojí.
V další části studie Snively a Theodor předkládají výsledky metody konečných prvků a opět srovnávají stegocerase s různými sudokopytníky. Bylo zjištěno, že přítomnost silné vrstvy rohoviny na vrchu stegocerasovy kupole by dokázala rozptýlit tlak na velkou plochu a tím snížit jeho účinek. Větší nápor působí na houbovitý vnitřek kupole než na kompaktní vnějšek, i zde je ale zátěž minimální. Ani v těch nejřidších místech kupole není tlak dostatečný k rozdrcení kosti. Při simulovaném nárazu o síle 1360 newtonů činí součinitel bezpečnosti stále 5 až 10. Největší tlak, který během testů působil na špičku kupole, byl 46 MPa. Rohovinová čepička vedla k jeho prudkému snížení na 8 MPa. Tlaky působící pod mozkovnou jsou větší než ty, které působí nad ní. Stejně jako u sudokopytníků, nesou i u stegocerase největší zátěž týlní kondyly. Houbovitá kost uvnitř kupole nicméně působí jako polštář a v kombinaci s krčními svaly snižuje tlak působící na týlní hrbol a dno mozkovny na přijatelnou míru.
U chocholatky, jejíž vyklenuté čelo je daleko tenčí než kupole stegocerase, je tlak působící na místa pod bodem nárazu větší. Zvláště pak je vyšší tlak působící na mozkovnu seshora a celkový tlak na kosti kolem mozkovny. Na lebku pižmoně působí při nárazu dokonce ještě vyšší tlaky, ale zátěž se směrem hlouběji rychleji snižuje. Na houbovitou kost nad mozkovnou a v týlních kondylech působí větší zátěž, než na kompaktní kost vršku lebky, přesto je však namáhání vrchní strany mozkovny menší, než u všech ostatních trkajících sudokopytníků. Růžky žirafy se podle modelů musejí při nárazech vypořádat s ještě vyššími tlaky, než rohy pižmoně. Na kompaktní kost nad mozkovnou sice působí jen nízký tlak i zátěž, obě hodnoty lze ale podstatně zvýšit zasažením prostředního růžku místo všech tří najednou. Snively & Theodor (2011) nicméně sami uznávají, že nárazová síla, s níž při modelech počítali, byla nereálně velká. Z dalších taxonů si dovolím vynechat vidloroha a zmíním pouze lamu, kde tlak – i přes umělé zesílení lebky pro účely analýzy dosahoval rekordních hodnot 60 MPa při rozloženém nárazu a dokonce 120 MPa při nárazu na hřeben temenní kosti, což už by kost rozštíplo. Způsobeno je to tenkou lebeční klenbou v kombinaci s řídkou kostí; při skutečném nárazu by navíc působil ještě vyšší tlak.
Co se týče výsledků rekurzivního dělení, nejlepší je zřejmě jejich prezentace formou tabulky:

Rekurzivní dělení ukázalo, že pravděpodobnost, že se Stegoceras účastnil nějakého druhu trkání, je při jeho zahrnutí mezi recentní zvířata značná.
S potvrzením této základní hypotézy se Snively a Theodor začínají věnovat tomu, o jaký druh chování konkrétně šlo. U různých sudokopytníků jsou adaptace na srážky hlavami různé, a vymřelý pachycefalosaur mnoho z nich kombinoval. Nejbližšími ekvivalenty dinosaura se zdají být chocholatka a poté ovce tlustorohá, a to i přesto, že disponují rohy a lebečními dutinami, které u stegocerase nenajdeme. Ovce tlustorohé se srážejí nejen svými rohy, ale i vrškem hlavy, což je chování blízké tomu, jaké bylo v minulosti navrhováno pro pachycefalosaury. Autoři zmiňují, že to, co ukázala metoda konečných prvků na jejich kombinovaném data setu není odlišné od toho, co už dříve odhalili Farke (2008) zvlášť pro sudokopytníky a Snively & Cox (2008) zvlášť pro pachycefalosaury. Protože velký kostěný dóm stegocerase při analýze rozložil sílu účinněji, než nižší čelo chocholatky, a protože chocholatka zase měla navrch nad lamou s tenkými lebečními kostmi, zdá se, že tlusté a nijak neodlehčené čelní kosti jsou pro trkání mimořádně vhodné. Aby se kost, z níž je složená stegocerasova kupole, pod zátěží a tlakem rozlomila, bylo by potřeba pětkrát až desetkrát větší síly, než jakou simuloval nynější test. Závažné poškození se zdá být nepravděpodobné i pro mladé pachycefalosaury, jejichž kupole se ještě skládala hlavně z houbovité tkáně s rozptýlenými trámečky, budící dojem křehkosti. U dospělců se silnou vrstvou kompaktní kosti by pak tlak a zátěž působící na mozkovnu byly zcela zanedbatelné.
Schopnost rozptýlit sílu nárazu se zdá být u stegocerase ještě větší, než u kteréhokoli ze zkoumaných sudokopytníků, kteří přitom prokazatelně trkání praktikují. Na druhou stranu nižší míra vaskularizace kupole u starších jedinců, kterou poprvé zpozorovali Goodwin & Horner (2004) a která je znovu demonstrována níže, svědčí o neschopnosti vyživovat případnou rohovinovou čepičku a nižší schopnosti hojení, která by mohla pro "trkací hypotézu" představovat závažný problém. Autoři přenechávají tento problém budoucím studiím a zmiňují, že relevantní se může ukázat především srovnání histologie a tomografických dat od stegocerase a chocholatky, která trká i navzdory absenci rohovinové ochranné vrstvy. Tuto výhradu ovšem Snively & Theodor (2011) považují za slabou oproti evidenci pro trkání, kterou se jim povedlo nashromáždit: stejně jako trkaví sudokopytníci, má i Stegoceras pod místem případného nárazu silnou vrstvu houbovité tkáně; trubičky kompaktní kosti, v nichž byly uloženy cévy, jsou analogií kostěných vzpěr v lebečních dutinách žijících taxonů; a neurovaskulární kanály ústící na povrch lebky jsou korelátem rohovinových čepiček (Hieronymus et al. 2009), kryjících hlavy trkavých sudokopytníků a zoborožců. Do budoucna také doporučují nasazení přechodové analýzy (transient analysis) a experimenty s rozptylem energie. Metoda konečných prvků totiž simulovala impakt staticky a lineárně, a přestože by skutečná simulace kolize zřejmě hodnoty tlaku a zátěže příliš nezměnila, mohly by lépe započítat efekt trámečků v houbovité kosti při rozptylu energie. Kostěné vzpěry, jaké v kupoli stegocerase formují trubičky okolo neurovaskulárních kanálů, zřejmě absorbují zátěž lépe než vzpěry, které by byly rovnoběžné se silou dopadu. I s dosavadními daty je však hypotéza o trkání silně podpořena, a my máme zpátky jednu z elegantních hypotéz dinosauří renezance, dosud pokládanou za vyvrácenou ošklivými fakty.
2. Schott et al. (2011) o stegocerasově ontogenezi: Ornatotholus = Stegoceras
Druhá studie se zabývá pachycefalosauřím růstem, který má závažné důsledky pro alfa taxonomii, tedy rozeznávání jednotlivých terminálních kladů ("druhů"). Je zde totiž důvodné podezření, že pachycefalosauří ontogeneze byla doprovázena velkými změnami ve tvaru lebky. I ten nejcharakterističtější znak skupiny, frontoparietální kupole, by mohl být ontogeneticky proměnlivý, což s sebou mnoho problémů. Mnoho klasifikací pachycefalosaurů totiž tuto skupinu dělilo na taxony s plochým vrškem lebky (tzv. homalocefalidy) a na ty s kupolí (pachycefalosauridy). Staré klasifikace (např. Sues & Galton 1987) pokládaly obě skupiny za monofyletické, novější fylogenetické analýzy ale tuto hypotézu nepodporují a bývalé "homalocefalidy" ukazují jako parafyletickou sérii taxonů čím dál tím bližších monofyletickým pachycefalosauridům (např. Schott et al. 2009).
Historicky byla poměrně oblíbená hypotéza, že pachycefalosauři užívali zesílené lebeční kosti k podobným soubojům, jako dnešní ovce tlustorohé (Ovis canadensis) nebo antilopy chocholatky, které sdílí s pachycefalosaury velmi tlusté, zaoblené kosti čelní. Dinosauří renezance 70. let, v níž paleontologové poprvé po dlouhé době začali o dinosaurech uvažovat jako o aktivních zvířatech, podobných spíše dnešním savcům než krokodýlům, rychle přijala hypotézu o trkání za svou. Sues (1978) např. demonstroval její biomechanickou přijatelnost na plexisklových modelech. Jenže jako tomu bylo i s jinými, často až příliš "heretickými" myšlenkami dinosauří renezance, ani představa pachycefalosaurů svádějících souboje prudkými údery hlavou o hlavu nepřečkala důkladnější rozbor. Zamítla ji histologická studie Goodwina a Hornera (2004), podle níž takovou činnost nedovolovala struktura kupole, v níž se pod povrchovou vrstvou kompaktní kosti skrývají sloupcovité trámečky (trabeculae) kolmé na vnější povrch dómu. Jiné práce ale tento výsledek zpochybnily: Snively & Cox (2008) argumentovali, že pokud by kostěnou kupoli pokrývala dostatečně silná vrstva rohoviny, nemožné by to být nemuselo; a trámcovitá struktura mohla být elastičtější, než si Goodwin a Horner mysleli. Farke (2008) zase s využitím metody konečných prvků ukázal, že lebeční dutiny u kozy domácí (Capra hircus) nezpůsobují snížení mechanické zátěže při nárazu, jak se dosud myslelo – ba právě naopak, modely s kompaktními čelními kostmi vykazovaly menší zátěž, a modely s vyklenutými, tlustými čelními kostmi často dokonce ještě menší. Hypotéza, že lebeční dutiny nebo kosti, které tyto dutiny ohraničují, dovedou absorbovat šok, tak byla jeho daty spíše vyvrácena. Absence vzduchem vyplněných prostor v lebečních kupolích pachycefalosaurů přitom byla v minulosti rovněž použita proti hypotéze o trkání (Fastovsky & Weishampel 2005).
Snively & Theodor (2011) se proto nyní pokoušejí dobrat se z těchto vzájemně rozporných indicií konečné odpovědi. Jejich studie aplikuje CT sken a 3D analýzu metodou konečných prvků na taxony, od nichž trkání hlavami prokazatelně známe (sudokopytníci chocholatka bělobřichá [Cephalophus leucogaster], pižmoň severní [Ovibos moschatus] a žirafa [Giraffa camelopardalis]), a na taxon, který je z trkání podezřelý – pachycefalosaur Stegoceras validum. Jelikož se ale žijící savci mlátí hlavami velmi vynalézavými způsoby, zahrnuli do analýzu taky ovci tlustorohou, která se vyhýbá kontaktu "roh na roh"; vidloroha amerického (Antilocapra americana), který naopak využívá své rohy k přetlačování a povrchem hlavy netrká; a také wapitiho (Cervus canadensis), pekari páskovaného (Tayasu tajacu) a lamu krotkou (Lama glama), kteří nedělají ani jedno.
Autoři dosti originálně nevyužívají data získaná od sudokopytníků k predikcím o stegocerasovi, ale data o stavbě stegocerasovy lebky k predikcím o stavbě lebek sudokopytníků. Konkrétně Snively a Theodor stavějí na tom, že v lebeční kupoli pachycefalosaurů (označované též přesnějším pojmem frontoparietální, který říká, že se skládá z kostí čelních [frontálů] a temenních [parietálů]) najdeme hluboko pod vrstvou kompaktní hutné kosti houbovitou kostní tkáň, a očekávají, že u turovitých bychom měli nalézt stejné uspořádání. Dále testují, zda kombinace lebečních dutin a hutné kosti umožňuje účinnější rozptyl tlaku, zda (a případně jak) by různě tvarované rohovinové polštáře v tomto ohledu pomohly, a zda bude stejný náraz na lebky stegocerase a žijících savců působit stejnou mechanickou námahou. Využívají k tomu algoritmus rekurzivního dělení (recursive partitioning analysis), který je znám především svým uplatněním medicíně: umožňuje stanovit diagnózu na základě známých korelací mezi symptomy a nemocemi. Snively & Theodor (2011) jej využívají přesně analogicky, když chtějí diagnostikovat chování zvířete z jeho morfologických znaků.

Hustota vnitřku kostěné kupole stegocerase, zjištěná pomocí počítačové tomografie exempláře UA 2. Obrázky A až F zachycují CT skeny šesti různě umístěných příčných průřezů lebky ze šikmého pohledu zepředu. Modrá znamená nejnižší hustotu, červená nejvyšší. Obrázek lebky z bočního pohledu zcela nahoře zachycuje jednotlivé roviny průřezu. Hustota a tloušťka kompaktní kosti roste směrem ke špičce kupole jak na předozadní (B–D), tak i pravolevé (C, D) ose. Kostěné trámečky vybíhají přibližně kolmo na vnější povrch kupole, jak je patrné na modré (a tím pádem řídké) oblasti na obrázcích B až E. Extrémní hustota povrchové kompaktní kosti může být CT skenem poněkud přehnaná, tlustá a hustá vrstva kompaktní kosti zde ale každopádně je. Trubičky kompaktní kosti, v nichž zřejmě byly uloženy cévy a které vybíhají až na vnější povrch kupole, jsou vyznačeny bílými čarami na obrázcích D až E a mohly by být analogické ke kostěným vzpěrám v lebkách trkavých sudokopytníků. (Zdroj: Snively & Theodor 2011: Figure 2)
 Srovnání hustoty lebek stegocerase a žijící chocholatky bělobřiché (Cephalophus leucogaster). (A–C) podélné průřezy lebkou stegocerase, vedené rovinami znázorněnými na obrázku nad nimi. (D–F) podélné průřezy lebkou chocholatky, vedené opět výše ukázanými rovinami. Obrázek F zachycuje, že ve vyklenuté čelní kosti chocholatky se kompaktní a houbovitá kost vrství stejným způsobem, jako u pachycefalosaurovy kupole na obrázku B. Zkratky: (ec) mozkovna. Modrá znamená nejnižší hustotu, červená nejvyšší. (Zdroj: Snively & Theodor 2011: Figure 5)
Srovnání hustoty lebek stegocerase a žijící chocholatky bělobřiché (Cephalophus leucogaster). (A–C) podélné průřezy lebkou stegocerase, vedené rovinami znázorněnými na obrázku nad nimi. (D–F) podélné průřezy lebkou chocholatky, vedené opět výše ukázanými rovinami. Obrázek F zachycuje, že ve vyklenuté čelní kosti chocholatky se kompaktní a houbovitá kost vrství stejným způsobem, jako u pachycefalosaurovy kupole na obrázku B. Zkratky: (ec) mozkovna. Modrá znamená nejnižší hustotu, červená nejvyšší. (Zdroj: Snively & Theodor 2011: Figure 5)

CT sken odhalil, že frontoparietální kupole stegocerase se skládá z kosti s vysokou a rovnoměrnou hustotou, větší než je hustota kostěného patra nebo týlu. To částečně souhlasí s daty získanými od žijící chocholatky, kde je vnější povrch "kupole", tvořené vyklenutou částí čelních kostí, také hustší než okolí. U chocholatky ale dosahují srovnatelné hustoty také nosní kosti a boční část kostí temenních. Proximální (povrchu lebky bližší) část kostěných jader rohů je zde daleko hustší než rohovinová vrstva, která jádra obaluje. U pižmoně je tomu naopak: rohovina je hustší než houbovitá kost, z níž je tvořena lebka.
Pokud se ale zaměříme na vnitřek kosti a ne jen její povrch, zjistíme, že mezi dvěma vrstvami velmi husté a kompaktní kosti (histologické zóny 1 a 3 podle Goodwina a Hornera [2004]) se nachází méně hustý prostředek (zóna 2). Svrchní zóna kompaktní kosti je tlustší na špičce kupole než po jejích okrajích. Dutiny, v nichž se za života zvířete nacházely cévy, se sice nacházejí ve vrstvě houbovité kosti, jsou však ohraničeny kostí kompaktní, ačkoli méně hustou, než je ta na povrchu kupole. Tato kompaktní kost tedy formuje trubice, které ústí na vnějším povrchu kupole. Podobně hustá kost rovněž tvoří hluboko uvnitř kupole jakési "proužky" (patrné na obrázku výše na snímcích C a D), které jsou zřejmě pozůstatkem raných stadií růstu kupole (což je téma, které je podrobně rozebráno níže). Rozložení různě hustých kostí u stegocerase kopíruje to, které známe od chocholatky. I tam najdeme hustou, kompaktní vrstvu přilehlou k mozkovně, nad kterou se klene nejprve houbovitá a blízko povrchu opět velice hustá kompaktní kostní tkáň. Velikost a hustota kostních trámečků je u chocholatky rozmanitější než u stegocerase: zatímco u vymřelého pachycefalosaura vybíhají kolmo k vnějšímu povrchu, u žijícího sudokopytníka vytvářejí houbovité mřížoví jakýchsi podpěr. Rovněž u ovce tlustorohé vidíme vrstvy v pořadí hluboká kompaktní, střední houbovitá a povrchová kompaktní. Pro pižmoně toto uspořádání platí v zásadě také (zaměříme-li se na relevantní část lebky, což v tomto případě není vyklenuté čelo, ale zkroucené rohy), chybí mu ale spodní vrstva kompaktní mozkovny obklopující mozkovnu. Ta je místo toho ohraničena houbovitou kostí, která pokračuje skrze většinu délky rohů až po špičku, kde je vystřídána vrstvou kompaktní kosti, překrytou následně ještě rohovinovou pochvou. Tato vrstva je tenčí než u stegocerase. Na rozdíl od tohoto pachycefalosaura také hustota povrchové vrstvy čelních kostí pižmoně nepřevyšuje hustotu nasálů a kostí horní čelisti.
U dalších sudokopytníků se lebeční dutiny, houbovitá kost a kompaktní kost vyskytují v nejrůznějších kombinacích. Žirafa má vrstvu houbovité kosti podstatně tenčí a objem dutin nad mozkem podstatně větší, než ovce tlustorohá. V jejích růžcích (osikonech) nicméně najdeme povrchovou vrstvu velmi husté kompaktní kosti a hluboko položenou vrstvu stejné, avšak o něco řidší kostní tkáně právě tak, jak je tomu u stegocerase. Pekari má nad mozkovnou dutiny, ale ne houbovitou kost, a wapiti, lama i vidloroh postrádají obojí.
V další části studie Snively a Theodor předkládají výsledky metody konečných prvků a opět srovnávají stegocerase s různými sudokopytníky. Bylo zjištěno, že přítomnost silné vrstvy rohoviny na vrchu stegocerasovy kupole by dokázala rozptýlit tlak na velkou plochu a tím snížit jeho účinek. Větší nápor působí na houbovitý vnitřek kupole než na kompaktní vnějšek, i zde je ale zátěž minimální. Ani v těch nejřidších místech kupole není tlak dostatečný k rozdrcení kosti. Při simulovaném nárazu o síle 1360 newtonů činí součinitel bezpečnosti stále 5 až 10. Největší tlak, který během testů působil na špičku kupole, byl 46 MPa. Rohovinová čepička vedla k jeho prudkému snížení na 8 MPa. Tlaky působící pod mozkovnou jsou větší než ty, které působí nad ní. Stejně jako u sudokopytníků, nesou i u stegocerase největší zátěž týlní kondyly. Houbovitá kost uvnitř kupole nicméně působí jako polštář a v kombinaci s krčními svaly snižuje tlak působící na týlní hrbol a dno mozkovny na přijatelnou míru.
U chocholatky, jejíž vyklenuté čelo je daleko tenčí než kupole stegocerase, je tlak působící na místa pod bodem nárazu větší. Zvláště pak je vyšší tlak působící na mozkovnu seshora a celkový tlak na kosti kolem mozkovny. Na lebku pižmoně působí při nárazu dokonce ještě vyšší tlaky, ale zátěž se směrem hlouběji rychleji snižuje. Na houbovitou kost nad mozkovnou a v týlních kondylech působí větší zátěž, než na kompaktní kost vršku lebky, přesto je však namáhání vrchní strany mozkovny menší, než u všech ostatních trkajících sudokopytníků. Růžky žirafy se podle modelů musejí při nárazech vypořádat s ještě vyššími tlaky, než rohy pižmoně. Na kompaktní kost nad mozkovnou sice působí jen nízký tlak i zátěž, obě hodnoty lze ale podstatně zvýšit zasažením prostředního růžku místo všech tří najednou. Snively & Theodor (2011) nicméně sami uznávají, že nárazová síla, s níž při modelech počítali, byla nereálně velká. Z dalších taxonů si dovolím vynechat vidloroha a zmíním pouze lamu, kde tlak – i přes umělé zesílení lebky pro účely analýzy dosahoval rekordních hodnot 60 MPa při rozloženém nárazu a dokonce 120 MPa při nárazu na hřeben temenní kosti, což už by kost rozštíplo. Způsobeno je to tenkou lebeční klenbou v kombinaci s řídkou kostí; při skutečném nárazu by navíc působil ještě vyšší tlak.
Co se týče výsledků rekurzivního dělení, nejlepší je zřejmě jejich prezentace formou tabulky:

Tabulka, prezentující data a současně výsledky metody rekurzivního dělení, použité zde k odvození typu chování (dole vpravo). Stegoceras, pižmoň, žirafa, lama, pekari, vidloroh, ovce, buvol, zoborožec štítnatý, kůň a chocholatka jsou (v pořadí odshora dolů) nejprve porovnáni z hlediska morfologie (morphological categories); konkrétně na přítomnost rozsáhlé vrstvy houbovité kosti (extensive trabeculae), tloušťku kompaktní kosti (layer thickness), její hustotu (density), přítomnost velkých kostěných vzpěr (large struts), přítomnost cév ustících na povrch lebky (surface vessels), tvar hlavy (head shape) a šířku krčních svalů (neck muscles). Následuje porovnání založené na datech z analýzy konečných prvků (biomechanical categories), které rozeznává jen dvě kategorie: zátěž lebky (braincase strain) a rozptyl tlaku (stress dissipation). Na základě toho je pomocí rekurzivního dělení předpovězeno, zda zvířata vykazují nějaké bojovné chování (agonism) a jaké by konkrétně mělo být: údery hlavou (head strikes), trkání (head butting) či žádné z výše uvedeného. ("Body blows ", čili nárazy tělem doplněné do kolonky k žirafě, zřejmě nejsou výsledkem analýzy, protože z morfologických dat týkajících se výhradně lebky mohly být jen těžko odvozeny.) Biomechanická data byla sice do rekurzivního dělení zahrnuta, jelikož ale nejsou známa ode všech zahrnutých taxonů, způsobila menší jednoznačnost výsledků. U žirafy je tloušťka kompaktní kosti měřena v rámci růžků. (Zdroj: Snively & Theodor 2011: Table 2)
Rekurzivní dělení ukázalo, že pravděpodobnost, že se Stegoceras účastnil nějakého druhu trkání, je při jeho zahrnutí mezi recentní zvířata značná.
S potvrzením této základní hypotézy se Snively a Theodor začínají věnovat tomu, o jaký druh chování konkrétně šlo. U různých sudokopytníků jsou adaptace na srážky hlavami různé, a vymřelý pachycefalosaur mnoho z nich kombinoval. Nejbližšími ekvivalenty dinosaura se zdají být chocholatka a poté ovce tlustorohá, a to i přesto, že disponují rohy a lebečními dutinami, které u stegocerase nenajdeme. Ovce tlustorohé se srážejí nejen svými rohy, ale i vrškem hlavy, což je chování blízké tomu, jaké bylo v minulosti navrhováno pro pachycefalosaury. Autoři zmiňují, že to, co ukázala metoda konečných prvků na jejich kombinovaném data setu není odlišné od toho, co už dříve odhalili Farke (2008) zvlášť pro sudokopytníky a Snively & Cox (2008) zvlášť pro pachycefalosaury. Protože velký kostěný dóm stegocerase při analýze rozložil sílu účinněji, než nižší čelo chocholatky, a protože chocholatka zase měla navrch nad lamou s tenkými lebečními kostmi, zdá se, že tlusté a nijak neodlehčené čelní kosti jsou pro trkání mimořádně vhodné. Aby se kost, z níž je složená stegocerasova kupole, pod zátěží a tlakem rozlomila, bylo by potřeba pětkrát až desetkrát větší síly, než jakou simuloval nynější test. Závažné poškození se zdá být nepravděpodobné i pro mladé pachycefalosaury, jejichž kupole se ještě skládala hlavně z houbovité tkáně s rozptýlenými trámečky, budící dojem křehkosti. U dospělců se silnou vrstvou kompaktní kosti by pak tlak a zátěž působící na mozkovnu byly zcela zanedbatelné.
Schopnost rozptýlit sílu nárazu se zdá být u stegocerase ještě větší, než u kteréhokoli ze zkoumaných sudokopytníků, kteří přitom prokazatelně trkání praktikují. Na druhou stranu nižší míra vaskularizace kupole u starších jedinců, kterou poprvé zpozorovali Goodwin & Horner (2004) a která je znovu demonstrována níže, svědčí o neschopnosti vyživovat případnou rohovinovou čepičku a nižší schopnosti hojení, která by mohla pro "trkací hypotézu" představovat závažný problém. Autoři přenechávají tento problém budoucím studiím a zmiňují, že relevantní se může ukázat především srovnání histologie a tomografických dat od stegocerase a chocholatky, která trká i navzdory absenci rohovinové ochranné vrstvy. Tuto výhradu ovšem Snively & Theodor (2011) považují za slabou oproti evidenci pro trkání, kterou se jim povedlo nashromáždit: stejně jako trkaví sudokopytníci, má i Stegoceras pod místem případného nárazu silnou vrstvu houbovité tkáně; trubičky kompaktní kosti, v nichž byly uloženy cévy, jsou analogií kostěných vzpěr v lebečních dutinách žijících taxonů; a neurovaskulární kanály ústící na povrch lebky jsou korelátem rohovinových čepiček (Hieronymus et al. 2009), kryjících hlavy trkavých sudokopytníků a zoborožců. Do budoucna také doporučují nasazení přechodové analýzy (transient analysis) a experimenty s rozptylem energie. Metoda konečných prvků totiž simulovala impakt staticky a lineárně, a přestože by skutečná simulace kolize zřejmě hodnoty tlaku a zátěže příliš nezměnila, mohly by lépe započítat efekt trámečků v houbovité kosti při rozptylu energie. Kostěné vzpěry, jaké v kupoli stegocerase formují trubičky okolo neurovaskulárních kanálů, zřejmě absorbují zátěž lépe než vzpěry, které by byly rovnoběžné se silou dopadu. I s dosavadními daty je však hypotéza o trkání silně podpořena, a my máme zpátky jednu z elegantních hypotéz dinosauří renezance, dosud pokládanou za vyvrácenou ošklivými fakty.

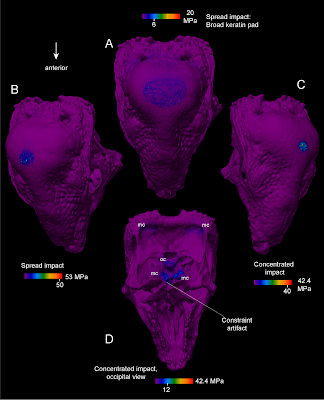
Efekt případné keratinové čepičky na tlaky, působící na stegocerasovu kupoli při trkání. Síla nárazu byla při modelování konstatně 1360 N. Sílu tlaku udává barevná škála. (A) Pohled na lebku seshora. Náraz je absorbován tlustou vrstvou rohoviny, u které dojde k deformaci; tlak je ale bezpečně rozložen na velkou plochu. Vrcholné zatížení v místě dopadu dosahuje 6 MPa, modální pak 3 MPa. (B) a (C) počítají s tenčí vrstvou rohoviny a tlak se tím stává soustředěnější. (D) pohled zezdola na stejný model, jaký je seshora vidět na obrázku (C). Vidět jsou mj. týlní kondyly (oc), kterými se lebka kloubila ke krční páteři. Pro detailnější popisky k obrázku viz původní studii. (Zdroj: Snively & Theodor 2011: Figure 13)
2. Schott et al. (2011) o stegocerasově ontogenezi: Ornatotholus = Stegoceras
Druhá studie se zabývá pachycefalosauřím růstem, který má závažné důsledky pro alfa taxonomii, tedy rozeznávání jednotlivých terminálních kladů ("druhů"). Je zde totiž důvodné podezření, že pachycefalosauří ontogeneze byla doprovázena velkými změnami ve tvaru lebky. I ten nejcharakterističtější znak skupiny, frontoparietální kupole, by mohl být ontogeneticky proměnlivý, což s sebou mnoho problémů. Mnoho klasifikací pachycefalosaurů totiž tuto skupinu dělilo na taxony s plochým vrškem lebky (tzv. homalocefalidy) a na ty s kupolí (pachycefalosauridy). Staré klasifikace (např. Sues & Galton 1987) pokládaly obě skupiny za monofyletické, novější fylogenetické analýzy ale tuto hypotézu nepodporují a bývalé "homalocefalidy" ukazují jako parafyletickou sérii taxonů čím dál tím bližších monofyletickým pachycefalosauridům (např. Schott et al. 2009).
A zde se znovu vracíme ke stegocerasovi. Jednou z prvních nalezených pachycefalosauřích lebek, která zřetelně vykazovala plochý vršek, byl exemplář s označením AMNH 5450. Galton (1971) jej popsal jako zástupce taxonu Stegoceras validum, a to konkrétně jako pohlavně dimorfickou samici. Po několika letech ovšem změnil názor a přesunul lebku do nového taxonu, Stegoceras browni (Wall & Galton 1979), pro který nakonec pocítil potřebu vztyčit nový "rod", Ornatotholus (Galton & Sues 1983). Součástí diagnózy nového taxonu přitom nebyla pouhá absence lebeční kupole, ale i větší průměr horního spánkového okna. Ukázalo se nicméně, že jak přítomnost lebeční kupole, tak i několik dalších, kriticky důležitých znaků (velká horní spánková okna a mělká příčná prohlubeň mezi vyvýšenými částmi čelních a temenních kostí) se u stegocerase během růstu měnila (Goodwin et al. 1998). Williamson & Carr (2002) na základě těchto poznatků navrhli, že by Ornatotholus mohl reprezentovat nedospělého jedince stegocerase. Sami ovšem věděli, že synonymizovat oba taxony by bylo problematické. Diagnóza stegocerase je totiž celá založená na znacích lebeční kupole, a protože tu Ornatotholus postrádá, nemohla být shoda obou taxonů nijak prokázána. Na Williamsonův a carrův krok různí autoři zareagovali různě. Zatímco někteří se s výše uvedeným problémem netrápili a oba taxony sesynonymizovali i tak, další si zase ornatotola ponechali, aniž by se snažili Williamsonovu a Carrovu hypotézu vyvrátit. To platí i poslední velké revizi pachycefalosaurů, za níž stáli Maryańska et al. (2004).
Situace se však od té doby ještě zkomplikovala. Williamsona a Carra trápilo, že neexistuje dobrý způsob, jak rozeznat plochohlavá mláďata odvozených pachycefalosaurů (disponujících v dospělosti lebeční kupolí) od pachycefalosaurů primitivních, plochohlavých ve všech fázích ontogeneze, jaké reprezentují např. Wannanosaurus nebo Homalocephale. V posledních letech ovšem začaly být tyto plochohlavé taxony jeden po jednom synonymizováni s tlustolebými taxony coby jejich mláďata. Horner & Goodwin (2009) argumentovali pro synonymitu plochohlavého drakorexe s tlustolebým pachycefalosaurem, Longrich et al. (2010) pro synonymitu plochohlavého homalocefaleho s kupoli-nesoucím taxonem Prenocephale. Ne všichni autoři ale s těmito kroky souhlasili: Evans et al. (2011) např. uznali, že od homalocefaleho známe pouze juvenilní jedince, nesouhlasili ovšem s tím, že by tato mláďata patřila prenocefalemu; a Sullivan (2007) navrhl, že u pachycefalosauridů mohlo docházet k pedomorfii, tedy k jevu, kdy si dospělí jedinci uchovávají znaky mláďat. Z toho je patrné, že ontogeneze pachycefalosaurů je komplikovaným tématem, jehož rozlousknutí může být přitom zásadní pro vyřešení mnoha taxonomických problémů, které zase mají velkou relevanci při výzkumu dávné biodiverzity.
Mnoho výše citovaných autorů – Evans, Horner, Goodwin, Longrich a Schott – nyní spojilo své síly, aby s pomocí srovnávací morfologie a počítačové tomografie ukázali, jak se měnila lebka pachycefalosauridů v průběhu ontogeneze. Výsledný model růstu a lebeční variability je dosud nejpodrobnějším svého druhu a Stegoceras pro něj představuje ideální zdroj dat. Tento taxon je znám z mnoha exemplářů a součástí jeho diagnózy je také ornamentace kosti šupinové (skvamosálu), která je nezávislá na změnách tvaru frontoparietální kupole. Díky tomu je možné testovat hypotézu, podle níž plochohlavé exempláře typu ornatotola představují pouze ranou fázi ontogenetické série "kupolovitých" taxonů.

Základní metodou, kterou autoři zvolili, je osteohistologie, neboli výzkum mikrostruktury kosti. Zpravidla bývá aplikován na dlouhé kosti, které ovšem vznikají jiným způsobem, než ploché kosti lebky: zatímco u těch prvních osifikuje chrupavka, která slouží budoucí kosti jako forma a zároveň jako jakési lešení; u lebečních kostí jde o osifikaci dezmogenní (intramembránovou), kdy kost vzniká přímo z mezenchymu v podkožním vazivu. Histologické metody, umožňující odlišit kosti dospělců a mláďat, lze však naštěstí aplikovat na oba typy kostí. Shodným znakem by měla být např. nižší vaskularizace starší kosti, která hrála velkou úlohu i ve výše rozebírané studii o pachycefalosauřím agonistickém chování. Otázkou je, jakým způsobem získat histologické informace, aniž by byl fosilní materiál nevratně poškozen. Goodwin et al. (2004) použili počítačovou tomografii s vysokým rozlišením – už tehdy mimochodem právě na ornatotola a stegocerase – a stejnou metodu uplatňuje i nynější studie. Ta CT skeny lebečních kostí různých exemplářů stegocerase využívá k otestování hypotézy, podle níž vaskularizace kosti klesá s rostoucím věkem. Potvrdila-li by se tato hypotéza, velmi by to pomohlo s odlišováním ontogeneticky daných rozdílů od těch, které jsou skutečně (= fylogeneticky) informativní, a otevřela by se cesta k vyřešení mnoha zapeklitých problémů pachycefalosauří systematiky. Autoři ovšem narazili na metodologické problémy: míru vaskularizace je z pouhé kosti těžké určit. Kostěné kanály totiž krom cév, které Schotta a spol. zajímají, pojímaly i spoustu jiných tkání. Dřívější studie Goodwina a Hornera (2004) – ta samá, která je citována výše jako útok na známou hypotézu o pachycefalosauřím trkání – tuto skutečnost ignorovala. Jak ale objasňují autoři, ani v jednom případě to moc nevadí. Goodwinovi a Hornerovi totiž šlo právě o celkový objem dutin v kupoli, zatímco nynější studie se o poměr mezi vaskularizací a objemem kostěných kanálů nemusí starat, pokud zůstává během ontogeneze zvířete konstantní. (A není důvod myslet si, že nezůstává.)
Zkoumanými exempláři jsou TMP 84.5.1, zřejmě mládě nebo suabdultní jedinec stegocerase; ROM 53555, jeden z největších znamých stegocerasů (jehož dospělost nicméně není zcela jistá) a AMNH 5450, holotyp ornatotola. Při sběru dílčích dat ale byly využívány i širší vzorky: např. pro morfometrickou studii, měřící vzdálenosti mezi 18 předem vytyčenými homologickými "orientačními body" na čelních a temenních kostech, bylo využito celkem 40 (!) stegocerasích lebek, počítaje v to i AMNH 5450. Autoři provedli sérii porovnání, v nichž vždy srovnávali dva logaritmicky transformované údaje s využitím RMA (reduced major axis) regrese. Aby dále otestovali hypotézu o alometrickém růstu frontoparietální kupole (podle níž se tento útvar zvětšoval rychlejším tempem, než kterým zvíře rostlo) a aby zjistili, jaký vliv mají na pozici regresní přímky plochohlavé exempláře, vyloučili Schott et al. (2011) z analýzy nejprve všechny plochohlavé exempláře, ponechali v ní však jedince podezřívané z toho, že zachycují prostřední fázi v růstu kupole. Při druhém měření byly dokonce vyloučeny všechny lebky, jejichž kupole byla tenčí než u TMP 84.5.1 – nejmenšího exempláře s jistotou patřícího stegocerasovi, který už nese plně vyvinutou kupoli.

Z měření vyplynulo, že výška stegocerasí kupole vykazuje pozitivní alometrii vůči šířce frontoparietálu a délce čelních kostí (= tloušťka kupole rostla rychleji, než šířka a délka kostí, z nichž byla složena). Silné důkazy pro pozitivní alometrii zcela vyvracejí předchozí závěry, k nimž došli Chapman et al. (1981). Autoři objasňují, jaké metodologické nedostatky tuto studii provázely. Chapman a spol. analyzovali značně nesourodý vzorek exemplářů, a to jak z hlediska geologické (3 různá souvrství), tak i stratigrafické (časový rozdíl až 5 milionů let) a fylogenetické (4 různé taxony – Stegoceras, Prenocephale, Hanssuesia, Colepiocephale) příslušnosti. Smíchali tak dohromady rozdíly, které mohly indikovat různá pohlaví, ontogenetická stadia nebo taxony. Jako základ studie si navíc nezvolili homologické orientační body, díky čemuž porovnávání hodnot naměřených u různých lebek ztratilo smysl. Šířka kupole už podle Schotta a spol. žádnou jednoznačnou tendenci nevykazuje a zdá se být buďto izometrická vůči šířce frontoparietálu (= roste stejně rychle), nebo naopak negativně alometrická (= roste pomaleji). Autoři také zjistili, že vyloučení plochohlavých a ne-tak-úplně-kupolovitých exemplářů z analýzy způsobí menší sklon regresní přímky. Obecné trendy jsou stále patrné, zdají se ale být slabší a spíše než na alometrii ukazují na izometrii. Tloušťka frontoparietálu sice i nadále vykazuje pozitivní alometrii vůči jeho délce a šířce, ale rozdíly v rychlosti růstu zde nejsou statisticky významné a klidně by mohlo jít i o růst stejnoměrný (= izometrii). Autoři uvádějí několik možností, proč analýza s vyloučením plochohlavých exemplářů dává odlišné výsledky: navrhují, že by sklon regresní křivky mohly snižovat větší frontoparietály; dodávají ale, že jejich vzorek je příliš malý na to, aby šlo tuto hypotézu ověřit.
Na rozdíl od Chapmana a spol. se Schott s kolektivem nesetkali s žádnými známkami sexuálního dimorfizmu. Vzorek zkoumaných jedinců byl však poměrně malý a rozeznat v něm bimodální rozdělení tvaru kupole, které by pohlavní dvoutvárnost potvrdilo, by bylo obtížné. Pozorované odchylky v tvaru kupole lze lépe vysvětlit ontogenetickou než pohlavní proměnlivostí. Autoři navrhují, že výsledky Chapmana a spol. znovu ovlivnilo zahrnutí nesouvisejících taxonů a také široký časový interval, z nějž exempláře pocházely (díky kterému se např. mohly nakumulovat anagenetické změny, díky čemuž by mladší formy vypadaly jinak než ty starší). Konstatují ovšem, že současná data hypotézu o pohlavním dimorfizmu nevylučují: za předpokladu, že se obě pohlaví vyvíjela nyní navrženým způsobem (tj. od ploché hlavy ke kupolovité), je taková možnost plně slučitelná s nynějšími poznatky a do budoucna by stála za otestování.
Histologický průzkum "ornatotola" odhalil hluboké švy mezi čelními kostmi a také jejich vysokou vaskularizaci (dutiny zabíraly 20% z celkového objemu frontoparietálu). Stejně tak exemplář TMP 84.5.1 obsahuje vysoce vaskularizovanou tkáň (17%). TMP 84.5.1 je přitom pozoruhodný tím, že zatímco čelní kosti a přední část těch temenních už jsou jasnou součástí rostoucí kupole, zadní část parietálu je sice poměrně tlustá, ale do kupole se ještě nezapojila. V případě ROM 53555, která už má plně vyvinutou kupoli, tvoří ale celkový objem dutin jen 3% a u srovnatelně velké UALVP 2 je to kolem 7%. Hypotéza Goodwina a Hornera (2004) o souvislosti mezi mírou vaskularizace a vývojem frontoparietální kupole tedy byla potvrzena. Morfometrická data zase ukazují nejen, že tloušťka frontoparietálu rostla rychleji než jeho šířka a délka, ale také na to, že v místě švu mezi čelními a temenními kostmi se jeho šířka zvětšovala rychleji než nad očima, a že kost temenní rostla do délky rychleji než kost čelní. Tato fakta ukazují, že typická "hruškovitá" kupole stegocerase mohla vzniknout z víceméně ploché lebky, jakou pozorujeme o ornatotola. Schott et al. (2011) rovněž dodávají, že vývoj kupole ovlivňoval i jiné kosti než samotný frontoparietál. Výrazně se měnily okrajové části lebky, včetně typického lemu (shelf) vzadu a po stranách, na němž se podílí kosti šupinové, temenní, zaočnicové a nadočnicové. Tyto kosti byly postupně začleňovány do zdvihající se kupole, čímž se lem stával méně výrazným.
Jelikož neinvazivní analýza vaskularizace pomocí CT skenu dokázala určit stáří jedince stejně dobře, jako detailní morfometrie, navrhují autoři používat do budoucna právě tuto metodu k testování hypotéz o synonymitě. První ukázku nabízejí sami: Ornatotholus browni je synonymizován s taxonem Stegoceras validum kvůli absenci diagnostických znaků, které by nebyly známy jako ontogeneticky proměnlivé či přítomné u stegocerase. Zvláštní hrbolkovitá textura na svrchním povrchu čelních, temenních, nadočnicových, zaočnicových a šupinových kostí je přítomna nejen u UCMZ(VP) 2008.001 – lebky, která je uznávaná coby plochohlavý exemplář stegocerase –, ale i na některých místech kupole plně dospělých stegocerasů a celé řady dalších pachycefalosaurů (Homalocephale, Goyocephale, "Dracorex", "Stygimoloch"*). Galton & Sues (1983) zase do diagnózy ornatotola zahrnuli velká horní spánková okna, zřetelně rozsáhlejší než ta stegocerasova. Důvody, proč je tento znak nevhodný, jsou hned dva. Jednak se mohly tyto otvory s ontogenezí uzavírat, jak podotkli Williamson & Carr (2002), jednak jsou jejich rozměry mezi různými jednotlivci velmi proměnlivé: plochohlavý UCMZ(VP) 2008.001 má paradoxně horní spánkové okno téměř o pět milimetrů menší, než "kupolovitý" TMP 84.5.1. Exemplář CMN 138, který kupolí také disponuje, jej má zase o 2,5 mm menší. Schott et al. (2011) uzavírají tuto záležitost s tím, že význam daného znaku bude ještě nutné prostudovat. Konečně posledním údajným diagnostickým znakem "ornatotola" je přítomnost mělké, příčně orientované prohlubně mezi dvěma části nízké kupole. Tento znak není znám od žádného jiného exempláře stegocerase včetně těch plohohlavých, což z něj činí zřejmě nejvážnější argument pro případnou separaci "ornatotola". Schott et al. (2011) jej ale intepretují jako přechodnou fázi v růstu kupole, během níž čelní a temenní kosti "bobtnaly" nezávisle na sobě, aby se nakonec spojily do jediného útvaru.
*Taxony jsou v uvozovkách proto, že je Horner & Goodwin (2009) v jedné ze svých nejkontroverznějších "lumperských" studií oba sesynonymizovali s pachycefalosaurem jako jeho mladší jedince.

Autoři proto v sekci Systematická paleontologie provedli formální synonymizaci taxonů Stegoceras browni Wall & Galton 1979 a Ornatotholus browni Galton & Sues 1983 se Stegoceras validum, pro nějž poskytují novou diagnózu prostou ontogeneticky proměnlivých (a tím pádem taxonomicky neinformativních) znaků. Uvedené autapomorfie zahrnují výše zmíněnou hrbolkovitou ornamentaci na bočních a zadních stranách šupinových kostí (jak bylo uvedeno výše, stejná textura se vyskytuje i u jiných pachycefalosaurů, ale jinde); výraznou řadu 5 až 8 nahoru vybíhajících výčnělků na posterodorzálním okraji obou kostí šupinových; řadu drobnějších hrbolků ve tvaru kýlu na boční straně týchž kostí. Autoři pak autapomorfické (= jedinečné) znaky doplňují ještě diferenční diagnózou, sestávající ze znaků, které stegocerase odlišují pouze od ostatních pachycefalosaurů. Sem patří chybějící ornamentace nosní kosti; silně redukovaná mezera mezi zuby v horní čelisti, a co do (levopravé) šířky zploštělý; plátu podobný pubický pedunkl ilia (výběžek na kyčelní kosti pro skloubení s kostí stydkou); a lebeční kupole méně vyvinutá, než u jiných pachycefalosaurů, kteří jí disponují (Pachycephalosaurus, Prenocephale, Sphaerotholus).
Svou práci Schott et al. (2011) uzavírají konstatováním, že během stegocerasova růstu docházelo k drastickým změnám tvaru lebky, které jsou příčinou značně rozdílného vzhledu mláďat a dospělců. Tento zvláštní průběh ontogeneze zřejmě nebyl unikátní pro stegocerase – spíše šlo o běžnou pachycefalosauří vývojovou trajektorii. Nšlo by konec konců o nic neobvyklého, jelikož radikální morfologické změny doprovázející růst jsou známy i od jiných ptakopánvých dinosaurů, včetně hadrosauridů (Evans 2010) a rohatých ceratopsů (Horner & Goodwin 2006). Díky tomu byla mláďata a dospělci řazeni do různých taxonů, což vedlo k přehnaným odhadům biodiverzity.* Dodnes zůstává otázkou, jak extrémní podobu mohly ontogenetické změny některých taxonů mít: kontroverzní jsou zde především studie se jménem Jack Hornera, který nejprve pomocí komplikovaných modelů resorpce kostěných rohů synonymizoval tři taxony pachycefalosaurů (Horner & Goodwin 2009) a na základě důkladné analýzy kostěného (parietoskvamosálního) límce navrhl synonymitu torosaura a nedoceratopse se známějším triceratopsem (Scannella & Horner 2010, na blogu zde a zde). Schott a spol. poznamenávají, že jejich studie ukazuje, jak do budoucna v případě podobných problémů postupovat: důležitý je velký vzorek zkoumaných jedinců a integrativní přístup, kombinující více nezávislých druhů důkazů (morfometrie, histologie).
*Nesouvisející poznámka, kterou si ale nemohu odpustit: celý problém ukazuje, že odhad biodiverzity by se neměl dělat sčítáním taxonů o stejné "úrovni", ať už jsou to "čeledě", "rody" nebo "druhy". Existují metody, které míru biodiverzity vypočítají z délek větví nebo počtu uzlů na kladogramu; nejznámější je z nich asi Faithův Phylogenetic Diversity Index (Faith 1992).
Situace se však od té doby ještě zkomplikovala. Williamsona a Carra trápilo, že neexistuje dobrý způsob, jak rozeznat plochohlavá mláďata odvozených pachycefalosaurů (disponujících v dospělosti lebeční kupolí) od pachycefalosaurů primitivních, plochohlavých ve všech fázích ontogeneze, jaké reprezentují např. Wannanosaurus nebo Homalocephale. V posledních letech ovšem začaly být tyto plochohlavé taxony jeden po jednom synonymizováni s tlustolebými taxony coby jejich mláďata. Horner & Goodwin (2009) argumentovali pro synonymitu plochohlavého drakorexe s tlustolebým pachycefalosaurem, Longrich et al. (2010) pro synonymitu plochohlavého homalocefaleho s kupoli-nesoucím taxonem Prenocephale. Ne všichni autoři ale s těmito kroky souhlasili: Evans et al. (2011) např. uznali, že od homalocefaleho známe pouze juvenilní jedince, nesouhlasili ovšem s tím, že by tato mláďata patřila prenocefalemu; a Sullivan (2007) navrhl, že u pachycefalosauridů mohlo docházet k pedomorfii, tedy k jevu, kdy si dospělí jedinci uchovávají znaky mláďat. Z toho je patrné, že ontogeneze pachycefalosaurů je komplikovaným tématem, jehož rozlousknutí může být přitom zásadní pro vyřešení mnoha taxonomických problémů, které zase mají velkou relevanci při výzkumu dávné biodiverzity.
Mnoho výše citovaných autorů – Evans, Horner, Goodwin, Longrich a Schott – nyní spojilo své síly, aby s pomocí srovnávací morfologie a počítačové tomografie ukázali, jak se měnila lebka pachycefalosauridů v průběhu ontogeneze. Výsledný model růstu a lebeční variability je dosud nejpodrobnějším svého druhu a Stegoceras pro něj představuje ideální zdroj dat. Tento taxon je znám z mnoha exemplářů a součástí jeho diagnózy je také ornamentace kosti šupinové (skvamosálu), která je nezávislá na změnách tvaru frontoparietální kupole. Díky tomu je možné testovat hypotézu, podle níž plochohlavé exempláře typu ornatotola představují pouze ranou fázi ontogenetické série "kupolovitých" taxonů.

Holotyp "ornatotola" AMNH 5450 na snímcích z počítačové tomografie s vysokým rozlišením (HRCT) seshora (A vlevo), zboku (B vlevo), zepředu (C vlevo), v plošném průřezu (A vpravo), sagitálním průřezu (B vpravo) a příčném průřezu (C vpravo). Šipky ukazují na místa, kde CT sken nezachytil pravou strukturu kosti, ale mineralizovaná místa odpovídající kolagenovým vláknům, která procházela napříč a podél švu mezi čelními kostmi. Zkratky: (f) čelní kost, (f-f) šev mezi čelními kostmi, (f-p) šev mezi kostí čelní a temenní, (p) temenní kost, (Z-1 až Z-3) histologické zóny 1 až 3, zavedené Goodwinem a Hornerem (2004) a zmiňované rovněž výše. (Zdroj: Schott et al. 2011: Figure 7)
Základní metodou, kterou autoři zvolili, je osteohistologie, neboli výzkum mikrostruktury kosti. Zpravidla bývá aplikován na dlouhé kosti, které ovšem vznikají jiným způsobem, než ploché kosti lebky: zatímco u těch prvních osifikuje chrupavka, která slouží budoucí kosti jako forma a zároveň jako jakési lešení; u lebečních kostí jde o osifikaci dezmogenní (intramembránovou), kdy kost vzniká přímo z mezenchymu v podkožním vazivu. Histologické metody, umožňující odlišit kosti dospělců a mláďat, lze však naštěstí aplikovat na oba typy kostí. Shodným znakem by měla být např. nižší vaskularizace starší kosti, která hrála velkou úlohu i ve výše rozebírané studii o pachycefalosauřím agonistickém chování. Otázkou je, jakým způsobem získat histologické informace, aniž by byl fosilní materiál nevratně poškozen. Goodwin et al. (2004) použili počítačovou tomografii s vysokým rozlišením – už tehdy mimochodem právě na ornatotola a stegocerase – a stejnou metodu uplatňuje i nynější studie. Ta CT skeny lebečních kostí různých exemplářů stegocerase využívá k otestování hypotézy, podle níž vaskularizace kosti klesá s rostoucím věkem. Potvrdila-li by se tato hypotéza, velmi by to pomohlo s odlišováním ontogeneticky daných rozdílů od těch, které jsou skutečně (= fylogeneticky) informativní, a otevřela by se cesta k vyřešení mnoha zapeklitých problémů pachycefalosauří systematiky. Autoři ovšem narazili na metodologické problémy: míru vaskularizace je z pouhé kosti těžké určit. Kostěné kanály totiž krom cév, které Schotta a spol. zajímají, pojímaly i spoustu jiných tkání. Dřívější studie Goodwina a Hornera (2004) – ta samá, která je citována výše jako útok na známou hypotézu o pachycefalosauřím trkání – tuto skutečnost ignorovala. Jak ale objasňují autoři, ani v jednom případě to moc nevadí. Goodwinovi a Hornerovi totiž šlo právě o celkový objem dutin v kupoli, zatímco nynější studie se o poměr mezi vaskularizací a objemem kostěných kanálů nemusí starat, pokud zůstává během ontogeneze zvířete konstantní. (A není důvod myslet si, že nezůstává.)
Zkoumanými exempláři jsou TMP 84.5.1, zřejmě mládě nebo suabdultní jedinec stegocerase; ROM 53555, jeden z největších znamých stegocerasů (jehož dospělost nicméně není zcela jistá) a AMNH 5450, holotyp ornatotola. Při sběru dílčích dat ale byly využívány i širší vzorky: např. pro morfometrickou studii, měřící vzdálenosti mezi 18 předem vytyčenými homologickými "orientačními body" na čelních a temenních kostech, bylo využito celkem 40 (!) stegocerasích lebek, počítaje v to i AMNH 5450. Autoři provedli sérii porovnání, v nichž vždy srovnávali dva logaritmicky transformované údaje s využitím RMA (reduced major axis) regrese. Aby dále otestovali hypotézu o alometrickém růstu frontoparietální kupole (podle níž se tento útvar zvětšoval rychlejším tempem, než kterým zvíře rostlo) a aby zjistili, jaký vliv mají na pozici regresní přímky plochohlavé exempláře, vyloučili Schott et al. (2011) z analýzy nejprve všechny plochohlavé exempláře, ponechali v ní však jedince podezřívané z toho, že zachycují prostřední fázi v růstu kupole. Při druhém měření byly dokonce vyloučeny všechny lebky, jejichž kupole byla tenčí než u TMP 84.5.1 – nejmenšího exempláře s jistotou patřícího stegocerasovi, který už nese plně vyvinutou kupoli.

Růstová série stegocerase, sestavená z rozlišných plochohlavých i "kupolovitých" lebek, při pohledu seshora (nahoře) a zboku (dole). Zleva doprava je zachycen zrekonstruovaný průběh ontogeneze od nejmladších po nejstarší jedince. UCMZ(VP) 2008.001 začíná se zcela plochým frontoparietálem. Bývalý "Ornatotholus" (AMNH 5450) zachycuje stadium, kdy se kupole začíná zvedat oddělené na čelních a temenních kostech. ROM 53555 potom reprezentuje plně dospělého jedince s typickou hruškovitě tvarovanou kupolí, o něco méně vyvinutou než u jiných pachycefalosaurů. Poslední obrázek krom frontoparietálu zahrnuje i boční okraje lebky a (vpředu seříznutou) část nosních kostí. (Zdroj: Schott et al. 2011: Figure 4)
 Graf, zachycující vztahy mezi dvěma logaritmicky transformovanými proměnnými: šířkou frontoparietálu (osa x) a jeho tloušťkou (osa y). Regresní přímky jsou grafem proloženy metodou RMA (reduced major axis), přičemž červená platí pouze pro exempláře s vyvinutější kupolí (= tlustším frontoparietálem), než jakou nese TMP 84.5.1; modrá pro všechny předchozí exempláře + 3 lebky s neúplně vyvinutou kupolí; a šedá pro všechny exempláře včetně těch plochohlavých. Čísla podél os jsou logaritmy hodnot změřených v milimetrech. Kresba kostěné kupole dole ukazuje, v jakém místě byla měřena tloušťka frontoparietálu (T:fp). (Zdroj: Schott et al. 2011: Figure 6)
Graf, zachycující vztahy mezi dvěma logaritmicky transformovanými proměnnými: šířkou frontoparietálu (osa x) a jeho tloušťkou (osa y). Regresní přímky jsou grafem proloženy metodou RMA (reduced major axis), přičemž červená platí pouze pro exempláře s vyvinutější kupolí (= tlustším frontoparietálem), než jakou nese TMP 84.5.1; modrá pro všechny předchozí exempláře + 3 lebky s neúplně vyvinutou kupolí; a šedá pro všechny exempláře včetně těch plochohlavých. Čísla podél os jsou logaritmy hodnot změřených v milimetrech. Kresba kostěné kupole dole ukazuje, v jakém místě byla měřena tloušťka frontoparietálu (T:fp). (Zdroj: Schott et al. 2011: Figure 6)

Z měření vyplynulo, že výška stegocerasí kupole vykazuje pozitivní alometrii vůči šířce frontoparietálu a délce čelních kostí (= tloušťka kupole rostla rychleji, než šířka a délka kostí, z nichž byla složena). Silné důkazy pro pozitivní alometrii zcela vyvracejí předchozí závěry, k nimž došli Chapman et al. (1981). Autoři objasňují, jaké metodologické nedostatky tuto studii provázely. Chapman a spol. analyzovali značně nesourodý vzorek exemplářů, a to jak z hlediska geologické (3 různá souvrství), tak i stratigrafické (časový rozdíl až 5 milionů let) a fylogenetické (4 různé taxony – Stegoceras, Prenocephale, Hanssuesia, Colepiocephale) příslušnosti. Smíchali tak dohromady rozdíly, které mohly indikovat různá pohlaví, ontogenetická stadia nebo taxony. Jako základ studie si navíc nezvolili homologické orientační body, díky čemuž porovnávání hodnot naměřených u různých lebek ztratilo smysl. Šířka kupole už podle Schotta a spol. žádnou jednoznačnou tendenci nevykazuje a zdá se být buďto izometrická vůči šířce frontoparietálu (= roste stejně rychle), nebo naopak negativně alometrická (= roste pomaleji). Autoři také zjistili, že vyloučení plochohlavých a ne-tak-úplně-kupolovitých exemplářů z analýzy způsobí menší sklon regresní přímky. Obecné trendy jsou stále patrné, zdají se ale být slabší a spíše než na alometrii ukazují na izometrii. Tloušťka frontoparietálu sice i nadále vykazuje pozitivní alometrii vůči jeho délce a šířce, ale rozdíly v rychlosti růstu zde nejsou statisticky významné a klidně by mohlo jít i o růst stejnoměrný (= izometrii). Autoři uvádějí několik možností, proč analýza s vyloučením plochohlavých exemplářů dává odlišné výsledky: navrhují, že by sklon regresní křivky mohly snižovat větší frontoparietály; dodávají ale, že jejich vzorek je příliš malý na to, aby šlo tuto hypotézu ověřit.
Na rozdíl od Chapmana a spol. se Schott s kolektivem nesetkali s žádnými známkami sexuálního dimorfizmu. Vzorek zkoumaných jedinců byl však poměrně malý a rozeznat v něm bimodální rozdělení tvaru kupole, které by pohlavní dvoutvárnost potvrdilo, by bylo obtížné. Pozorované odchylky v tvaru kupole lze lépe vysvětlit ontogenetickou než pohlavní proměnlivostí. Autoři navrhují, že výsledky Chapmana a spol. znovu ovlivnilo zahrnutí nesouvisejících taxonů a také široký časový interval, z nějž exempláře pocházely (díky kterému se např. mohly nakumulovat anagenetické změny, díky čemuž by mladší formy vypadaly jinak než ty starší). Konstatují ovšem, že současná data hypotézu o pohlavním dimorfizmu nevylučují: za předpokladu, že se obě pohlaví vyvíjela nyní navrženým způsobem (tj. od ploché hlavy ke kupolovité), je taková možnost plně slučitelná s nynějšími poznatky a do budoucna by stála za otestování.
Histologický průzkum "ornatotola" odhalil hluboké švy mezi čelními kostmi a také jejich vysokou vaskularizaci (dutiny zabíraly 20% z celkového objemu frontoparietálu). Stejně tak exemplář TMP 84.5.1 obsahuje vysoce vaskularizovanou tkáň (17%). TMP 84.5.1 je přitom pozoruhodný tím, že zatímco čelní kosti a přední část těch temenních už jsou jasnou součástí rostoucí kupole, zadní část parietálu je sice poměrně tlustá, ale do kupole se ještě nezapojila. V případě ROM 53555, která už má plně vyvinutou kupoli, tvoří ale celkový objem dutin jen 3% a u srovnatelně velké UALVP 2 je to kolem 7%. Hypotéza Goodwina a Hornera (2004) o souvislosti mezi mírou vaskularizace a vývojem frontoparietální kupole tedy byla potvrzena. Morfometrická data zase ukazují nejen, že tloušťka frontoparietálu rostla rychleji než jeho šířka a délka, ale také na to, že v místě švu mezi čelními a temenními kostmi se jeho šířka zvětšovala rychleji než nad očima, a že kost temenní rostla do délky rychleji než kost čelní. Tato fakta ukazují, že typická "hruškovitá" kupole stegocerase mohla vzniknout z víceméně ploché lebky, jakou pozorujeme o ornatotola. Schott et al. (2011) rovněž dodávají, že vývoj kupole ovlivňoval i jiné kosti než samotný frontoparietál. Výrazně se měnily okrajové části lebky, včetně typického lemu (shelf) vzadu a po stranách, na němž se podílí kosti šupinové, temenní, zaočnicové a nadočnicové. Tyto kosti byly postupně začleňovány do zdvihající se kupole, čímž se lem stával méně výrazným.
Jelikož neinvazivní analýza vaskularizace pomocí CT skenu dokázala určit stáří jedince stejně dobře, jako detailní morfometrie, navrhují autoři používat do budoucna právě tuto metodu k testování hypotéz o synonymitě. První ukázku nabízejí sami: Ornatotholus browni je synonymizován s taxonem Stegoceras validum kvůli absenci diagnostických znaků, které by nebyly známy jako ontogeneticky proměnlivé či přítomné u stegocerase. Zvláštní hrbolkovitá textura na svrchním povrchu čelních, temenních, nadočnicových, zaočnicových a šupinových kostí je přítomna nejen u UCMZ(VP) 2008.001 – lebky, která je uznávaná coby plochohlavý exemplář stegocerase –, ale i na některých místech kupole plně dospělých stegocerasů a celé řady dalších pachycefalosaurů (Homalocephale, Goyocephale, "Dracorex", "Stygimoloch"*). Galton & Sues (1983) zase do diagnózy ornatotola zahrnuli velká horní spánková okna, zřetelně rozsáhlejší než ta stegocerasova. Důvody, proč je tento znak nevhodný, jsou hned dva. Jednak se mohly tyto otvory s ontogenezí uzavírat, jak podotkli Williamson & Carr (2002), jednak jsou jejich rozměry mezi různými jednotlivci velmi proměnlivé: plochohlavý UCMZ(VP) 2008.001 má paradoxně horní spánkové okno téměř o pět milimetrů menší, než "kupolovitý" TMP 84.5.1. Exemplář CMN 138, který kupolí také disponuje, jej má zase o 2,5 mm menší. Schott et al. (2011) uzavírají tuto záležitost s tím, že význam daného znaku bude ještě nutné prostudovat. Konečně posledním údajným diagnostickým znakem "ornatotola" je přítomnost mělké, příčně orientované prohlubně mezi dvěma části nízké kupole. Tento znak není znám od žádného jiného exempláře stegocerase včetně těch plohohlavých, což z něj činí zřejmě nejvážnější argument pro případnou separaci "ornatotola". Schott et al. (2011) jej ale intepretují jako přechodnou fázi v růstu kupole, během níž čelní a temenní kosti "bobtnaly" nezávisle na sobě, aby se nakonec spojily do jediného útvaru.
*Taxony jsou v uvozovkách proto, že je Horner & Goodwin (2009) v jedné ze svých nejkontroverznějších "lumperských" studií oba sesynonymizovali s pachycefalosaurem jako jeho mladší jedince.

Obrázek znázorňující, jak Schott a spol. došli ke svým odhadům vaskularizace. V horní řadě jsou zachyceny lebky AMNH 5450 (vlevo), TMP 84.5.1 (uprostřed) a ROM 53555 (vpravo), v prostřední řadě CT skeny jejich relevantních částí a v dolní řadě příčný průřez čelní kostí, vedený rovinou vyznačenou na obrázcích v horní řadě červenou čarou. V místě zájmu (vyznačeném červeným obdélníkem) byl změřen podíl plochy dutin na celkové ploše průřezu, který reprezentuje míru vaskularizace. Výsledky v procentech jsou připsané k obrázkům. (Zdroj: Schott et al. 2011: Figure 10)
Autoři proto v sekci Systematická paleontologie provedli formální synonymizaci taxonů Stegoceras browni Wall & Galton 1979 a Ornatotholus browni Galton & Sues 1983 se Stegoceras validum, pro nějž poskytují novou diagnózu prostou ontogeneticky proměnlivých (a tím pádem taxonomicky neinformativních) znaků. Uvedené autapomorfie zahrnují výše zmíněnou hrbolkovitou ornamentaci na bočních a zadních stranách šupinových kostí (jak bylo uvedeno výše, stejná textura se vyskytuje i u jiných pachycefalosaurů, ale jinde); výraznou řadu 5 až 8 nahoru vybíhajících výčnělků na posterodorzálním okraji obou kostí šupinových; řadu drobnějších hrbolků ve tvaru kýlu na boční straně týchž kostí. Autoři pak autapomorfické (= jedinečné) znaky doplňují ještě diferenční diagnózou, sestávající ze znaků, které stegocerase odlišují pouze od ostatních pachycefalosaurů. Sem patří chybějící ornamentace nosní kosti; silně redukovaná mezera mezi zuby v horní čelisti, a co do (levopravé) šířky zploštělý; plátu podobný pubický pedunkl ilia (výběžek na kyčelní kosti pro skloubení s kostí stydkou); a lebeční kupole méně vyvinutá, než u jiných pachycefalosaurů, kteří jí disponují (Pachycephalosaurus, Prenocephale, Sphaerotholus).
Svou práci Schott et al. (2011) uzavírají konstatováním, že během stegocerasova růstu docházelo k drastickým změnám tvaru lebky, které jsou příčinou značně rozdílného vzhledu mláďat a dospělců. Tento zvláštní průběh ontogeneze zřejmě nebyl unikátní pro stegocerase – spíše šlo o běžnou pachycefalosauří vývojovou trajektorii. Nšlo by konec konců o nic neobvyklého, jelikož radikální morfologické změny doprovázející růst jsou známy i od jiných ptakopánvých dinosaurů, včetně hadrosauridů (Evans 2010) a rohatých ceratopsů (Horner & Goodwin 2006). Díky tomu byla mláďata a dospělci řazeni do různých taxonů, což vedlo k přehnaným odhadům biodiverzity.* Dodnes zůstává otázkou, jak extrémní podobu mohly ontogenetické změny některých taxonů mít: kontroverzní jsou zde především studie se jménem Jack Hornera, který nejprve pomocí komplikovaných modelů resorpce kostěných rohů synonymizoval tři taxony pachycefalosaurů (Horner & Goodwin 2009) a na základě důkladné analýzy kostěného (parietoskvamosálního) límce navrhl synonymitu torosaura a nedoceratopse se známějším triceratopsem (Scannella & Horner 2010, na blogu zde a zde). Schott a spol. poznamenávají, že jejich studie ukazuje, jak do budoucna v případě podobných problémů postupovat: důležitý je velký vzorek zkoumaných jedinců a integrativní přístup, kombinující více nezávislých druhů důkazů (morfometrie, histologie).
*Nesouvisející poznámka, kterou si ale nemohu odpustit: celý problém ukazuje, že odhad biodiverzity by se neměl dělat sčítáním taxonů o stejné "úrovni", ať už jsou to "čeledě", "rody" nebo "druhy". Existují metody, které míru biodiverzity vypočítají z délek větví nebo počtu uzlů na kladogramu; nejznámější je z nich asi Faithův Phylogenetic Diversity Index (Faith 1992).
Zdroje:
- Chapman RE, Galton PM, Sepkoski JJ Jr, Wall WP 1981 A morphometric study of the cranium of the pachycephalosaurid dinosaur Stegoceras. J Paleont 55(3): 608–18
- Evans DC 2010 Cranial anatomy and ontogeny of Hypacrosaurus altispinus, and a comparative analysis of skull growth in lambeosaurines (Ornithischia: Hadrosauridae). Zool J Linn Soc 159(2): 398–434
- Evans DC, Brown CM, Ryan MJ, Tsogtbaatar K 2011 Cranial ornamentation and ontogenetic status of Homalocephalae calathocercos (Ornithischia: Pachycephalosauria) from the Nemegt Formation, Mongolia. J Vert Paleont 31: 84–92
- Faith DP 1992 Conservation evaluation and phylogenetic diversity. Biol Cons 61: 1–10
- Farke AA 2008 Frontal sinuses and head-butting in goats: a finite element analysis. J Exp Biol 211: 3085–94
- Fastovsky DF, Weishampel DB 2005 The Evolution and Extinction of the Dinosaurs. Cambridge, UK: Cambridge Univ Press
- Galton PM 1971 A primitive dome-headed dinosaur (Ornithischia: Pachycephalosauridae) from the Lower Cretaceous of England and the function of the dome of pachycephalosaurids. J Paleont 45(1): 40–7
- Galton PM, Sues H-D 1983 New data on pachycephalosaurid dinosaurs (Reptilia: Ornithischia) from North America. Can J Earth Sci 20: 462–72
- Goodwin MB, Buchholtz EA, Johnson RE 1998 Cranial anatomy and diagnosis of Stygimoloch spinifer (Ornithischia: Pachycephalosauria) with comments on cranial display structures in agonistic behavior. J Vert Paleont 18(2): 363–75
- Goodwin MB, Colbert M, Rowe T 2004 High-resolution computed tomography of the type Ornatotholus browni (Ornithischia: Pachycephalosauria) confirms its early ontogenetic stage and synonomy with Stegoceras. J Vert Paleont 24(3): 65A
- Goodwin MB, Horner JR 2004 Cranial histology of pachycephalosaurs (Ornithischia: Marginocephalia) reveals transitory structures inconsistent with head-butting behavior. Paleobiol 30: 253–67
- Hieronymus TL, Witmer LM, Tanke DH, Currie PJ 2009 The facial integument of centrosaurine ceratopsids: morphological and histological correlates of novel skin structures. Anat Rec 292: 1370–96
- Horner JR, Goodwin MB 2006 Major cranial changes in Triceratops. Proc R Soc B 273(1602): 2757–61
- Horner JR, Goodwin MB 2009 Extreme cranial ontogeny in the Upper Cretaceous dinosaur Pachycephalosaurus. PLoS ONE 4(10): e7626
- Longrich NR, Sankey J, Tanke D 2010 Texacephale langstoni, a new genus of pachycephalosaurid (Dinosauria: Ornithischia) from the upper Campanian Aguja Formation, southern Texas, USA. Cretac Res 31:274–284
- Maryańska T, Chapman RE, Weishampel DB 2004 Pachycephalosauria. 464–77 in Weishampel DB, Dodson P, Osmólska H, eds. The Dinosauria (2nd ed). Berkeley, CA: Univ of California Press
- Scannella JB, Horner JR 2010 Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny. J Vert Paleont 30(4): 1157–68
- Schott RK, Evans DC, Goodwin MB, Horner JR, Brown CM, Longrich NR 2011 Cranial ontogeny in Stegoceras validum (Dinosauria: Pachycephalosauria): a quantitative model of pachycephalosaur dome growth and variation. PLoS ONE 6(6): e21092
- Schott RK, Evans DC, Williamson TE, Carr TD, Goodwin MB 2009 The anatomy and systematics of Colepiocephale lambei (Dinosauria: Pachycephalosauridae). J Vert Paleont 29(3): 771–86
- Snively E, Cox A 2008 Structural mechanics of pachycephalosaur crania permitted head-butting behavior. Palaeont Electr 11(1):3A: 1–17
- Snively E, Theodor JM 2011 Common functional correlates of head-strike behavior in the pachycephalosaur Stegoceras validum (Ornithischia, Dinosauria) and combative artiodactyls. PLoS ONE 6(6): e21422
- Sues H-D 1978 Functional morphology of the dome in pachycephalosaurid dinosaurs. N Jahrb Geol Paläont Mh 8: 459–72
- Sues H-D, Galton PM 1987 Anatomy and classification of the North American Pachycephalosauria (Dinosauria: Ornithschia). Palaeont Abt A 198: 1–40
- Sullivan RM 2007 Doming, heterochrony, and paedomorphosis in the Pachycephalosauridae (Ornithischia: Dinosauria): taxonomic and phylogenetic implications. J Vert Paleont 27(Suppl. to 3): 154–5A
- Wall WP, Galton PM 1979 Notes on pachycephalosaurid dinosaurs (Reptilia: Ornithischia) from North America, with comments on their status as ornithopods. Can J Earth Sci 16: 1176–86
- Williamson TE, Carr TD 2002 A new genus of derived pachycephalosaurian from western North America. J Vert Paleont 22(4): 779–801
















